
PYTHONINAE Fitzinger, 1826
|
publication ID |
https://doi.org/ 10.5281/zenodo.4650515 |
|
persistent identifier |
https://treatment.plazi.org/id/EC1B3736-FFDE-8930-FF5B-FBB9CBD2FA16 |
|
treatment provided by |
Felipe |
|
scientific name |
PYTHONINAE Fitzinger, 1826 |
| status |
|
Subfamily PYTHONINAE Fitzinger, 1826
Genus Python Daudin, 1803
Python sp.
“unnamed species of Python ” – Rage in Thomas et al. 1982: 117.
“assez gros Boïné” – Rage in Demarcq et al. 1983: 256.
Python sp. – Ivanov 1997a: 35-36, fig. 18.
MATERIAL EXAMINED. — 1 palatine ( FSL 368001), 37 cervical vertebrae ( MNHN; VCO 5- VCO 9; FSL 368002- FSL 368032, FSL 368033), 321 trunk vertebrae ( MNHN; VCO 10- VCO 29; FSL 368034- 368334), 2 caudal vertebrae ( FSL 368335, FSL 368336).
DESCRIPTION
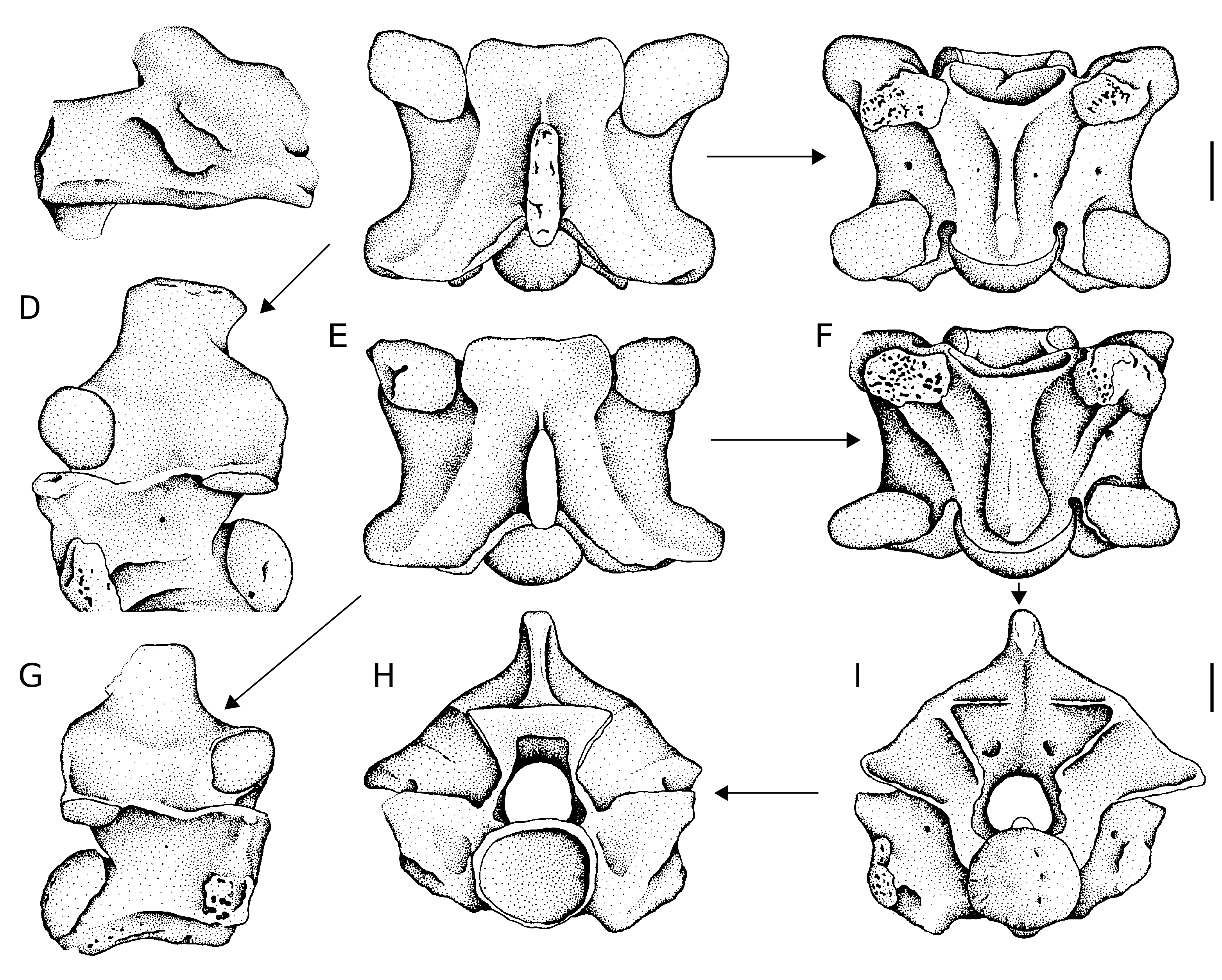
Palatine ( Fig. 2A View FIG )
The complete maxillary process and the base of the vomerine process are preserved. Both the rostral and the caudal parts of the bone are broken off. The maxillary process is relatively short, a distinct foramen for maxillary nerve penetrates its base. The vomerine process was directed anteriorly, however its distal end is broken off. The teeth are not preserved, with the exception of the base of one tooth.
Cervical vertebrae ( Fig. 2 View FIG B-D)
The vertebrae are massively built. In lateral view, the neural spine is as long as high or somewhat higher. The anterior cervical vertebrae possess caudally shifted neural spine; both the cranial and caudal margins of the neural spine are inclined posteriorly. In posterior cervical vertebrae, the cranial margin of the neural spine is almost vertical. The interzygapophyseal ridges are considerably developed and they are anteroventrally directed. Generally, the lateral foramina are very small and indistinct. The hypapophyses are broken off nearby their base. The subcentral ridges are arched slightly dorsally, in some cases they are straight. The indistinctly divided paradiapophyses are mostly heavy damaged. In dorsal view, the zygosphenal lip is almost straight with indistinct median lobe, or it is even slightly concave. The neural spine can be slightly thickened postero-dorsally, however its cranial margin is always sharp. The prezygapophyseal articular surfaces are irregular – originally, they could be subtriangular. In ventral view, both the subcentral ridges and grooves are mostly distinct, especially in the cranial part of vertebrae. The subcentral foramina are distinct. The postzygapophyseal articular surfaces are shaped irregularly. In cranial view, the neural arch is considerably arched, the neural canal is suborbicular. The zygosphenal lip is straight to slightly convex dorsally. The parapophyseal processes are short and rounded ventrally. The cotyle is rounded to slightly flattened dorso-ventrally. The paracotylar foramina are lacking.
Trunk vertebrae ( Fig. 2 View FIG E-I)
All vertebrae are damaged. In lateral view, the neural spines are broken off mostly at the base. The cranial margin of the neural spine is almost vertical, rarely it overhangs anteriorly. The caudal margin overhangs posteriorly. The sharp interzygapophyseal ridges are markedly directed anteroventrally. The lateral foramina are distinct, they are not situated in any depressions. The aliform processes can project behind the postzygapophyseal articular surfaces. Paradiapophyses are indistinctly divided. The subcentral ridges are arched dorsally. In dorsal view, the neural spine is thick, especially its caudal part. The cranial margin is sharp. The prezygapophyseal articular surfaces are subtriangular. The zygosphenal lip is wide with variable shape – it may be straight with welldeveloped lateral lobes, but sometimes the wide median lobe is present. The prezygapophyseal processes are very short. In ventral view, the haemal keel is wide, especially in large vertebrae. In some cases, a cranial expansion of the haemal keel is observed just behind the cotyle. The haemal keel is better developed in posterior trunk vertebrae where it becomes more pronounced because of deeper subcentral grooves and more distinct subcentral ridges. The subcentral ridges extend from the parapophyses to almost the base of the condyle. The subcentral foramina are well- perceptible. Laterally expanded postzygapophyseal articular surfaces are irregular. In cranial view, the margin of the massive zygosphene is straight, rarely concave. The neural arch is vaulted, the neural canal is about subtriangular with distinct lateral sinuses. The prezygapophyses are slightly turned up. The cotyle is almost rounded, rarely flattened dorso-ventrally. The paracotylar foramina are missing. In caudal view, is seen the massively build zygantrum. Metrical measurements are as follows (n = 24): cl: or = 6.04-10.11; naw: or = 7.08-13.42; cl/naw: or = 0.71-0.89, mean 0.79 + 0.04.
Caudal vertebrae
Both the pleurapophyses and haemapophyses are broken off at the base. In lateral view, the neural spine is short, somewhat longer than high. The interzygapophyseal ridges are distinct, the lateral foramina are very small. In dorsal view, the zygosphenal lip is straight or concave, lateral lobes are turned up dorsally in cranial view. The prezygapophyseal articular surfaces are expanded antero-laterally. They are irregularly, subtriangularly shaped. In cranial view, the neural arch is considerably vaulted, the cotyle is flattened dorso-ventrally and the paracotylar foramina are lacking.
DISCUSSION
The palatine of representatives of the subfamily Pythoninae is characterised by the distinct foramen at the base of the maxillary process (Underwood 1967). The discovered palatine is very fragmentary, however, because of the dimensions of the bone it may be assumed that it likely belongs to an adult representative of the genus Python . The vertebrae are massively built with a very low ratio cl/naw, which is apparently lower than 1.00. Moreover, the paracotylar foramina are absent. Therefore, it can be assumed that the discovered vertebrae belong to the Boidae . The allocation to the subfamily Pythoninae and the genus Python is based on the presence of well-preserved palatine. The fossil material could be compared only with the recent species Python reticulatus (Schneider, 1801)
A B C
(ZZSiD 436 juv., ZZSiD 437 juv.) and Python molurus (Linnaeus, 1758) (ZZSiD 460) – however, because of the scantiness of cranial bones a more precise determination was not possible. The vertebrae of Python sp. differ from the morphotypes Boidae B & C (see below) in: 1) the much more vaulted neural arch; 2) the shape of the massive zygosphenal lip.
At present, the available material represents the only known representative of the subfamily Pythoninae from the European Neogene. Another unquestionable Pythoninae comes from the German locality Messel (middle Eocene) where a great number of cranial bones, still articulated (including maxilla and palatine with taxonomically important features), have been discovered (Szyndlar & Böhme 1993). However, the axial skeleton was not precisely studied (Szyndlar & Böhme 1993). The extinct species Palaeopython sardus Portis, 1901 (=? Python sardus [Rage, 1984]) belongs most likely also to the subfamily Pythoninae . Both last mentioned constrictors were characterised by considerable diameters.
Subfamily ERYCINAE Bonaparte, 1831
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |