
Quedius subunicolor Korge, 1961
|
publication ID |
https://doi.org/ 10.37520/aemnp.2022.017 |
|
publication LSID |
lsid:zoobank.org:pub:28D55112-98B1-49A5-B382-58B1B068570B |
|
DOI |
https://doi.org/10.5281/zenodo.7503693 |
|
persistent identifier |
https://treatment.plazi.org/id/038987A0-FFDE-4B3C-B11A-A7781113FACA |
|
treatment provided by |
Felipe |
|
scientific name |
Quedius subunicolor Korge, 1961 |
| status |
|
Quedius subunicolor Korge, 1961 View in CoL
( Figs 1 View Fig , 2F View Fig , 4 View Fig , 7F View Fig , 13C View Fig , 20 View Fig )
Quedius subunicolor Korge, 1961: 81 View in CoL [Type locality: Schweden, Häggenås, Jämtland]
Quedius altaicus Korge, 1962: 152 View in CoL [Type locality: Zentral-Altai], syn. nov.
References. Pൺඅආ (1962): 187, (1963): 142 (characters); TංĸHඈආංඋඈඏൺ (1973): 189 (distribution); SHංඅඈඏ (1975): 376 (distribution); Cඈංൿൿൺංඍ (1978): 193 (characters); LඈHඌൾ & LඎർHඍ (1989): 172 (biology); Sൺඅ-ඇංඍඌĸൺ & Sඈඅඈൽඈඏඇංĸඈඏ (2018a): 128–130 (redescription, suspected synonym of Q. altaicus View in CoL ); (2019): 47 (key and characters).
Type material examined. Quedius subunicolor : Pൺඋൺඍඒඉൾඌ (MZLU):1J, labelled: “Häggenäs s-n Jtl.T. Palm 4–8, 8 1945 / det. H. Korge Quedius subunicolor Korge / Paratypus subunicolor Korge / Quedius subunicolor Korge / Type no. 1202:2 MZLU/ 2016 189 MZLU”; 3♀♀, same data, but last two labels “Type no: 1202:3 MZLU/ 2016 190 MZLU”.
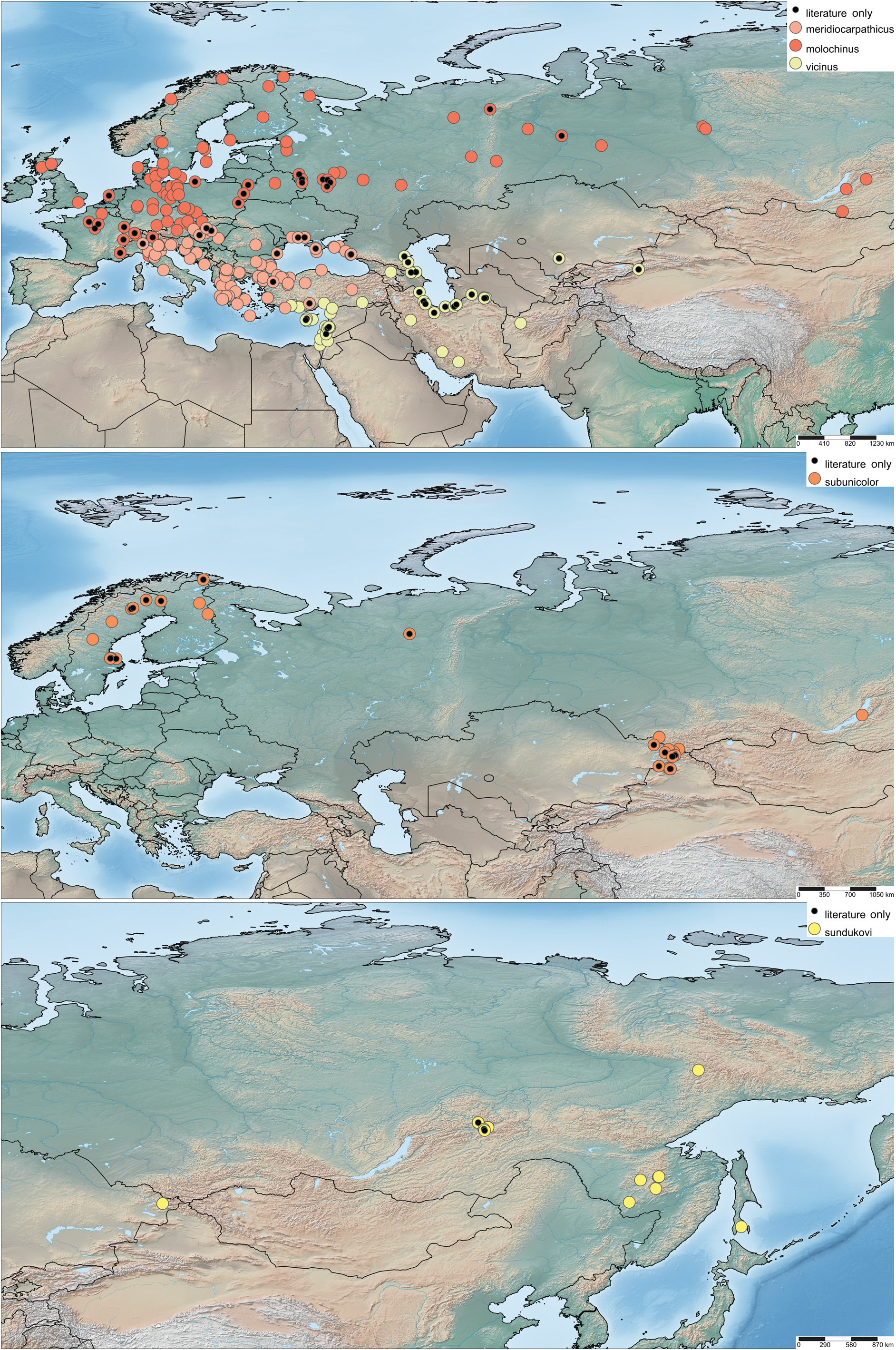
Quedius altaicus : Hඈඅඈඍඒඉൾ: ♀ (FMNH), labelled: “Zentral-Altai, lg. Leder, det.Bang-Haag / unicolor Kies. det.Bernhauer / ♀ Holotypus Quedius s. str. altaicus H. Korge / Chicago NHMus M. Bernhauer Collection / Holotype teste D.J. Clarke 2014 GDI Imaging Project / Photographed Kelsey Keaton 2014 Emu Catalog / FMNHINS 2819427 Field Museum”. Additional material examined. FINLAND: Kuusamo, [65.96, 29.17] (1 ♀ ZMUO); Savukoski, [67.29, 28.15], 15.-25.VI.2006, leg.M.Mutanen, P.Välimaki & M.Pentisaari (1J ZMUO). KAZAKHSTAN: Altai, Listvyaga, SSW Gora Tesninskiy Belok, Seredchikha River, [49.51, 85.25], 1200-1500 m, 27.VII.1997, R. Dudko & V. Zinchenko (1 J NHMD); Bukhtarma riv., Uryl-Chingistai, [49.19, 85.88], 13.VI.1987, leg. V.A. Kastcheev (1 J ZIN); Rahmanovskie Kluchi, [49.53, 86.50] 14.-16. VI.1980, leg.V.A. Kastcheev (1 J ZIN); Saur ridge, Uidene riv., [47.20, 85.32], 1600-1800 m, 15.VII.2011, leg.V.A. Kastcheev (10J 2♀♀ ZIN). RUSSIA: Aඅඍൺං Rൾඉ.: Source of Kuma river, 50.9989, 84.1647, 1479 m, mixed forest, under stones, 4.VII.2019, leg. A. Solodovnikov, A. K. Hansen, & M. J. Justesen (2 NHMD); Source of Kuma river, 50.9806, 84.1652, 1800 m, under stones, timber line, 3.VII.2019,A. Solodovnikov, A. K. Hansen, & M. J. Justesen (1 J NHMD). Rൾඉ. ඈൿ Bඎඋඒൺඍංൺ: Lake Baikal, Svjatoy Nos, [53.69, 108.86], 9.VIII.1997, leg. A. Shavrin (1 J NHMD). SWEDEN: Barsele, [65.04, 17.45], leg. Alan Dufberg (1 MZLU); Revsund, [62.89, 15.12], 27.V.1970, leg. R. Bergvall (3 MZLU).
Redescription. Measurements JJ (n = 6): HW = 1.67– 1.78 (1.74); HL = 1.42–1.56 (1.49); HL/HW 0.80–0.89 (0.86); PW = 2.24–2.40 (2.32); PL = 2.04–2.20 (2.12); PL/PW 0.90–0.96 (0.91); EW = 2.22–2.56 (2.46); EL = 2.29–2.47 (2.36); EL/EW 0.92–1.03 (0.96); EL/PL 1.06– 1.17 (1.11); PW/HW 1.50–1.64 (1.56); forebody length 5.78–6.04 (5.97). ♀♀ (n = 3): HW = 1.67–1.76 (1.73); HL = 1.44–1.49 (1.47); HL/HW 0.82–0.88 (0.85); PW = 2.13–2.33 (2.25); PL = 1.89–2.02 (1.97); PL/PW 0.83–0.94 (0.88); EW = 2.11–2.47 (2.30); EL = 2.00–2.36 (2.24); EL/EW 0.95–1.01 (0.97); EL/PL 1.00–1.25 (1.14); PW/ HW 1.45–1.62 (1.54); forebody length 5.47–5.82 (5.67).
Medium sized, robust species; body black ( Fig. 7F View Fig ).
Head black, distinctly transverse, with eyes medium sized (EyL/TL = 1.88–2.13 (2.02)); microsculpture of fine transverse waves, in some areas almost meshed; no interocular punctures between anterior frontal punctures (cf. Fig. 6F View Fig ); antennae brown with antennomeres 1–5 clearly darkened, all antennomeres elongate; palpi brown.
Thorax: pronotum black, slightly wider than long, wi-der than head, with microsculpture of transverse waves; three punctures in dorsal row and one to two in sublateral row with its posteriormost puncture reaching just beyond level of first puncture of dorsal row; scutellum punctured and pubescent; elytra black, uniformly pubescent, slightly wider than long, roughly as long as–slightly longer than pronotum; legs dark brown to black, tarsi usually lighter pale brown.
Abdomen black, tergites uniformly punctured, with slight iridescence.
Male. Aedeagus ( Fig. 13C View Fig ): paramere lanceolate with no or very slight medial attenuation, reaching just beyond apex of median lobe, with rather large sensory peg setae forming two long single rows fusing together towards apex; median lobe broad with gentle constriction to a point at apex, on parameral side with two small teeth pointing slightly basad, positioned at level near basal level of peg setae band of paramere; internal sac without a continuation of C-sclerite.
Differential diagnosis. Quedius subunicolor is very similar to the species of the molochinus -group with dark elytra – Q. unicolor , Q. sundukovi – and to the dark forms of Q. molochinus . It differs from Q. unicolor by the microsculpture of head consisting of transverse waves and rarely becoming meshed (clearly meshed frons appearing dull in Q. unicolor ). Also, genitalia are clearly different in both species, and they have completely non-overlapping distributions. Quedius subunicolor can be clearly distinguished from Q. sundukovi by the presence of the palisade fringe of tergite VII, which is missing in Q. sundukovi . It usually differs from the dark forms of Q. molochinus in the darker colored appendages and finer microsculpture. If in doubt the internal sacs are also clearly different in both species: in Q. subunicolor C-sclerite lacking an extension, which is present in Q. molochinus .
Synonymic notes. KඈඋGൾ (1961) described Quedius subunicolor from Sweden for fully darkened specimens that originally were assigned to Q. unicolor , a species which is actually restricted to the central and southern European mountains. Additional records showed that Q. subunicolor was rather widespread in Scandinavia (Pൺඅආ 1962, 1963; TංĸHඈආංඋඈඏൺ 1973). Nearly at the same time, KඈඋGൾ (1962) described Q. altaicus based on two females from an unspecified locality in ‘central Altai’. Sൺඅඇංඍඌĸൺ & Sඈඅඈൽඈඏඇංĸඈඏ (2018a) made a thorough morphological comparison between Q. altaicus and Q. subunicolor , including type material and demonstrated that the external characters claimed by KඈඋGൾ (1962) as unique for Q. altaicus compared with Q. subunicolor (microstructure of the head, proportions of the pronotum, chaetotaxy of the head and pronotum) vary continuously between both species. They suspected a synonymy and suggested that molecular comparisons of the two species should be conducted. The results of such an analysis here, that included newly collected material, suggest that specimens from Northern Europe and Altai are identical based on COI barcode (0.6% intraspecific variation among them in one clade; Table 2 View Table 2 ; Fig. 4 View Fig ). Since there is continuous morphological variation and no clear separation in COI, there is no evidence for separate species. We therefore propose that Quedius altaicus Korge, 1962 syn. nov. is placed in synonymy with Quedius subunicolor Korge, 1961 .
Quedius subunicolor has historically been confused with Q. unicolor . Many older specimens from Scandinavia and Northern Russia have been assigned to Q. unicolor , although these belong to Q. subunicolor . More recently, it has been erroneously reported from the Bohemian Forest in Central Europe, e.g., BඈHගඹ & Mൺඍෂඃටඹൾĸ (2004).
Bionomics. In Northern Europe Q. subunicolor is almost exclusively found in bogs and fens. This is a type of habitat that historically has often been drained for agriculture and peat gathering, thus the species may be rarer now than it used to be. In the Altai Mountains all hitherto known specimens were collected at the elevations 1200–2000 m. We (AKH and AS) have spent six days in the western Altai mountains in the early summer of 2019 searching for this species. There we sifted many types of debris, but without any luck. It was not until we started turning stones just below the timber line that we found the first specimen. In spite of six days of focused collecting, we encountered only three specimens, all collected from under rocks near streams in forested areas ( Fig. 2F View Fig ). The low number of specimens we encountered is in agreement with the very few numbers of specimens available in museums. The species is likely very rare and patchily distributed across its range. Contrary to the morphological and molecular homogeneity of this species, we see a clear difference in habitat preference between the Scandinavian populations restricted to bogs and Altaian populations confined to the banks of mountain creeks at higher altitudes. Such variation in habitat preference within the distribution range is not unique and is also found in other members of Quedius sensu stricto, e.g., Q. labradorensis . Presumably, these different habitats provide the same or similar parameters of temperatures and moisture for a species within a wide range.
Distribution. Quedius subunicolor currently displays a disjunct distribution between Northern Europe and Altai Mountains, with few specimens known from the Ukhta District in the Komi Region of Russia (SHංඅඈඏ 1975) and from Lake Baikal listed herein ( Fig. 20 View Fig ). It is still unknown if Q. subunicolor represents a species continuously distributed from Northern Europe to Altai, or if it is a hitherto unrecorded case of a boreo-montane disjunction between Northern Europe and Siberian mountains. Further sampling should be done in the areas of the apparently large distributional gaps.






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Quedius subunicolor Korge, 1961
| Hansen, Aslak Kappel, Brunke, Adam, Simonsen, Thomas & Solodovnikov, Alexey 2022 |
Quedius subunicolor
| KORGE H. 1961: 81 |