
Rhagasostoma mimosa ( Brydone, 1930 )
|
publication ID |
https://doi.org/ 10.5852/ejt.2018.490 |
|
publication LSID |
lsid:zoobank.org:pub:BE023137-CC5E-4DC5-94F6-B549BB140361 |
|
DOI |
https://doi.org/10.5281/zenodo.3849974 |
|
persistent identifier |
https://treatment.plazi.org/id/03D587D7-9647-FF99-F302-FA01FE02FDC8 |
|
treatment provided by |
Valdenar |
|
scientific name |
Rhagasostoma mimosa ( Brydone, 1930 ) |
| status |
|
Rhagasostoma mimosa ( Brydone, 1930)
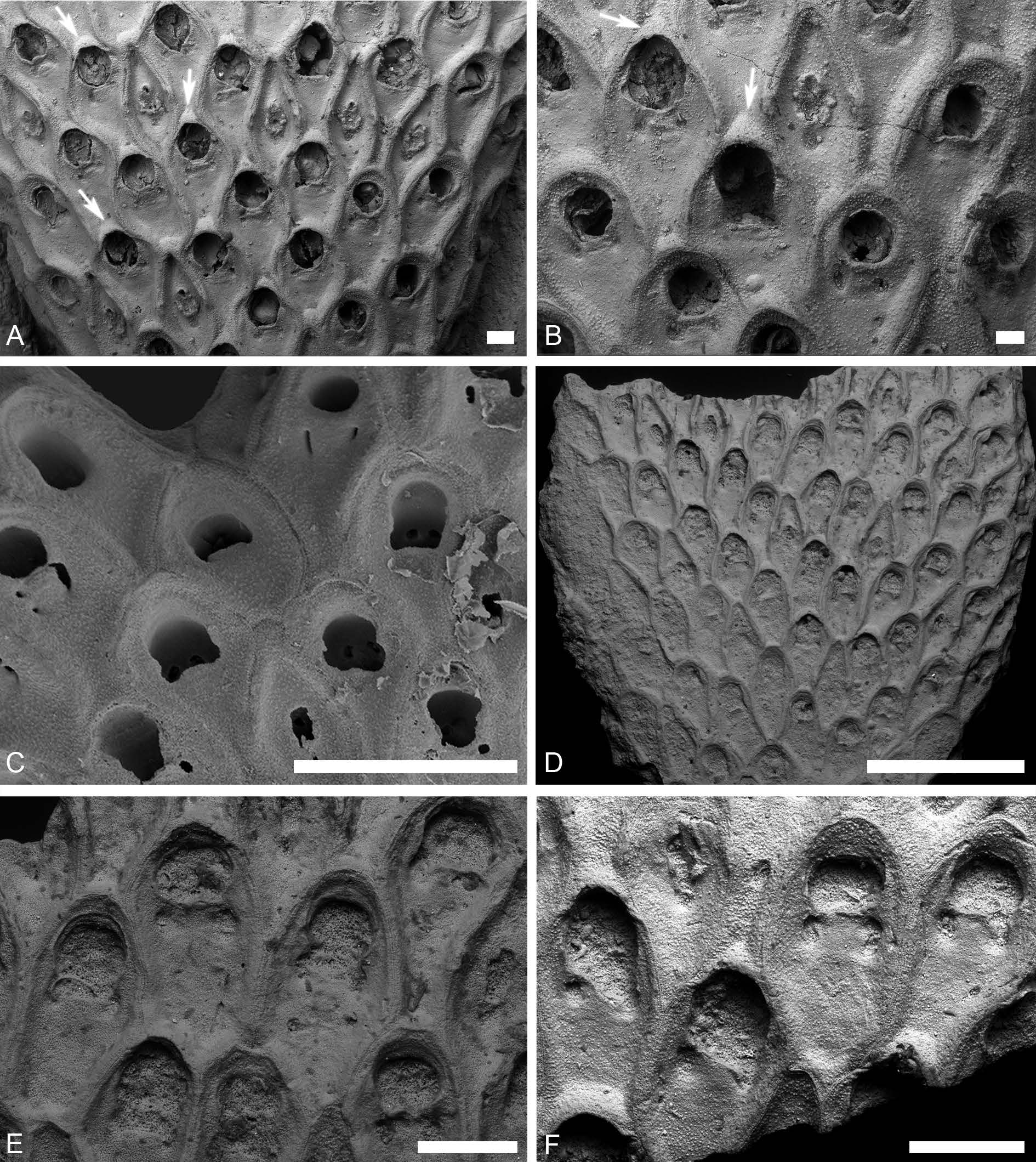
Fig. 19 View Fig , Table 11 View Table 11
Onychocella mimosa Brydone, 1930: 49 View in CoL , pl. 28, figs 1–2.
Woodipora mimosa – Voigt 1967: 50, pl. 18, fig. 4. — Schubert 1986: 40.
Woodipora rowei ( Brydone, 1906) – Schubert 1986 pars: 39, pl. 3, fig. 7, pl. 5, figs 4–5, pl. 8, figs 2–3. Onychocella mimosa View in CoL – Favorskaya 1992: 120 (mention). — Koromyslova 2014a: pl. 10, figs 1–2.
non Woodipora rowei – Schubert 1986 pars: 39, non pl. 3, figs 5–6, 8, pl. 5, figs 1–3, 6–8, pl. 8, fig. 1.
Material studied
Lectotype (here designated)
UNITED KINGDOM • England, Norfolk; Porosphaera Beds of Trimingham; early Maastrichtian ( Ostrea lunata Zone ); SM B36696 ( Fig. 19 View Fig AB; figured by Brydone 1930: pl. 28, figs 1–2).
Additional figured material
DENMARK • Island of Møn ; early Maastrichtian; SMF 29923 ( Fig. 19C View Fig ) .
KAZAKHSTAN • 2 specs; Mangystau Region;?early Maastrichtian of Hanga-Baba (урочиЩе Хангабаба) on the Mangyshlak Peninsula; PIN 5502/3056 ( Fig. 19F View Fig ), 5502/3057 ( Fig. 19 View Fig D–E).
Other material
BELARUS • 2 specs; Grodno Region; erratic block of?late Campanian age in a quarry near Hrodna / Grodno (Гродна/ Гродно); PIN 2922 View Materials /203, 2922/207 (imaged in Koromyslova 2014a: pl. 10, figs 1–2) .
Description
Colony rigidly erect, bifoliate, multiserial, branches 2.0–10.0 mm wide. Ancestrula and early astogeny not observed. Colony formed by pyriform and ovate autozooids and vicarious avicularia. Autozooids subrectangular with rounded distal ends; zooidal boundaries raised. Pyriform autozooids with a narrow proximal end but widening distally, the widening usually starting from the proximal margin of the opesia. In ovate autozooids, the proximal end is partly overlapped by the avicularian rostra and the autozooid begins to widen considerably below the proximal margin of the opesia, usually at the boundary with an avicularium. Gymnocyst lacking. Cryptocyst slightly granulated, slightly depressed centrally and occupying half or more of the autozooidal frontal surface, peripheral caverns lacking. Opesia terminal or subterminal. Opesial rim elevated, formed by a projecting, thickened cryptocyst, subcircular, a pair of opesiules located 70–90 µm proximallateral of opesia and divided from the latter by a cryptocystal tongue. Fusion of cryptocyst distal of opesiules and tongue of cryptocyst incomplete. Septula in walls not observed. Ovicells immersed, ooecium is formed by the distal zooid, triangular, vestigial, with slightly granulated surface, not protruding above the colony surface ( Fig. 19 View Fig A–B, D–F). Avicularia vicarious, rhomboidal. Rostrum conical in outline and symmetrical, usually with straight lateral walls; indentations or projections present at base of rostrum. Usually two types of avicularia differing mainly in the length of the rostrum and rostral apex, which partly overlaps the proximal end of the distal autozooids: avicularia with short rostra and short, trough-like apices (0–130 µm), and avicularia with long rostra without projecting apices. Proximal part short, narrowing downwards, with convex lateral sides. Entire frontal surface of avicularium occupied by a fragile, slightly granulated cryptocyst containing five openings: two small, subcircular openings, distally and proximally, in between three slit-like, parallel openings. Cryptocyst between openings frequently destroyed causing them to coalesce into a single opening of variable outline. Closure plates, kenozooids and intramural reparative budding in autozooids and avicularia not observed.
Remarks
Rhagasostoma mimosa was introduced by Brydone (1930) for species from the Porosphaera beds of Trimingham very much resembling R. rowei ( Brydone, 1906) but differing from the latter in having two opesiules. He considered the possibility that colonies of R. rowei were badly preserved samples of R. mimosa but concluded that the two must be different species because colonies of R. rowei are more slender and fragile, avicularia of R. mimosa have a central cavity enclosed by a marked wall, and R. rowei and R. mimosa never co-occur in the Chalk of England.
Voigt (1967) re-examined R. rowei and R. mimosa from Central Asia and rejected the morphological arguments used by Brydone (1930) to separate the two species. He concluded that except for R. mimosa having opesiules, the two species are completely identical, but as no intermediate stages occur, he regarded a separation of R. rowei and R. mimosa as justified. He argued that calcification of the frontal wall of R. rowei during the Late Cretaceous resulted over time in the formation of two opesiules completely separated from the opesia as passages for the bundles of parietal muscles. A similar development has also been observed in Rhagasostoma disparile (d’Orbigny, 1851) and Rhagasostoma strumulosum Marsson, 1887 (cf. Schubert 1986: fig. 12), which are indeed closely related to R. rowei and R. mimosa . Furthermore, Voigt (1967) assigned mimosa to the genus Woodipora Jullien, 1888 because of the opesiules, but kept rowei in Onychocella .
In his Woodipora monograph, Schubert (1986) regarded rowei and mimosa as two different phenotypes of the same species, which he assigned to Woodipora . The length and width measurements that he conducted on autozooids and avicularia and their opesiae revealed no significant distinction between rowei and mimosa . Furthermore, he argued that colonies in the mimosa stage often have autozooids with and also without opesiules (cf. Schubert 1986: pl. 8, fig. 3). As the zooids without opesiules are of primary origin and cannot have been destroyed during preparation or other processes, he concluded that the occurrence of opesiules is not species-specific and cannot be considered an argument for the existence of mimosa as an independent species. He regarded the mimosa stage as an ‘end-member’ of a parallel development by progressive calcification of the cryptocyst from the Coniacian to the Maastrichtian resulting in the formation of two opesiules. Zooids with advanced calcification of the cryptocyst, however, did not completely replace the rowei stage, the two stages thus co-occurring in single colonies.
Voigt (1991) again regarded mimosa and rowei (and also R. disparile and R. strumulosum ) as separate species. He saw the four species as an example of an intermediate status between onychocellid pseudomalacostegans and microporid or thalamoporellid coilostegans, placing disparile and rowei in Onychocella and strumulosum and mimosa in Woodipora . Thus, the microporid or thalamoporellid genus Woodipora would have originated several times by convergent evolution.
Without description or figures, Favorskaya (1992) mentioned Onychocella mimosa from the Maastrichtian of the southern Aral Sea Region in Uzbekistan and from Hanga-Baba (урочиЩе Ханга-баба), 30 km east of Fort Shevchenko (Форт-Шевченко) in the Mangystau Region, Kazakhstan.
A comprehensive description of morphological differences between R. rowei and R. mimosa from the Grodno quarry in Belarus was undertaken by Koromyslova (2014a). As she showed, the two species are not conspecific and differ in the shape of the autozooidal opesia and in the cryptocyst that develops opesiular indentations in R. rowei and opesiules in R. mimosa . Furthermore, specimens of R. rowei show intramural reparative budding in avicularia, which has not been observed in colonies of R. mimosa . Based on the length of the rostrum and the apex, three different types of avicularia have been observed in R. rowei , while R. mimosa shows only avicularia with short rostra and short trough-like apices ( Koromyslova 2014a). Avicularia of R. rowei and R. mimosa are indeed very distinct from most other species within the genus Rhagasostoma . Centrally located, there is an opesia and three opesiules in the avicularian cryptocyst.
We have restudied Brydone’s syntypes in the SM collection, choosing a lectotype (figured by Brydone 1930: pl. 28, figs 1–2), as well as the material from the collection of E. Voigt ( Fig. 18I View Fig , 19C View Fig ) and T.A. Favorskaya ( Fig. 19 View Fig D–F). The specimens labelled as Onychocella mimosa (SM B36697) from the Brydone Collection and (TsNIGR Museum 32/9757) from the Voigt Collection belong to R. rowei because the autozooids lack opesiules. As already discussed by Brydone (1930) and Voigt (1967), R. mimosa and R. rowei are very similar, differing only in R. mimosa having opesiae that are round instead of bell-shaped and opesiules instead of opesiular indentions. The presence of autozooids with opesiular indentations in colonies of R. mimosa in most cases can be explained by post-mortem destruction of the cryptocystal tongue that separated the opesiules from the opesia, or by the fact that the opesiae and opesiules of these autozooids were not formed fully and they were in the rowei stage (cf. Schubert 1986: pl. 8, fig. 3).
Distribution
Late Campanian Belarus: Quarry near Hrodna / Grodno (Гродна/ Гродно), Grodno Region ( Koromyslova 2014a). Early Maastrichtian Germany: Saturn quarry near Kronsmoor, Schleswig- Holstein ( Schubert 1986); glacial drift deposits in northern Germany ( Voigt 1967). Kazakhstan: unknown locality on the Mangyshlak Peninsula, Mangystau Region ( Favorskaya 1992). United Kingdom: Trimingham, Norfolk, England ( Brydone 1930). Uzbekistan: southern Aral Sea Region, Republic of Karakalpakstan ( Favorskaya 1992). Late Maastrichtian Denmark: Møns Klint, Sjaelland ( Schubert 1986). Turkmenistan: Gorge W of the “Kamyschli” gorge in the Kopet Dag Mountains NW of Aşgabat ( Voigt 1967).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SuperFamily |
Microporoidea |
|
Family |
|
|
Genus |
Rhagasostoma mimosa ( Brydone, 1930 )
| Koromyslova, Anna V., Taylor, Paul D., Martha, Silviu O. & Riley, Matthew 2018 |
Woodipora mimosa
| Schubert T. 1986: 40 |
| Voigt E. 1967: 50 |
Onychocella mimosa
| Brydone R. M. 1930: 49 |