
Rhyacophila aphrodite, Malicky, 1975
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4623.3.8 |
|
publication LSID |
lsid:zoobank.org:pub:6755FA6F-B5CE-4686-A317-1CBFEE0A19B0 |
|
persistent identifier |
https://treatment.plazi.org/id/E262D664-F47E-FFC4-8691-FDBEFE60FEE7 |
|
treatment provided by |
Plazi |
|
scientific name |
Rhyacophila aphrodite |
| status |
|
Morphological diagnosis of fifth instar larvae of Rhyacophila aphrodite View in CoL from those of other European Trichoptera
A summary of morphological features for the identification of European caddisfly families was given by Waringer & Graf (2013). Due to the mid-abdominal, tetrafilament gill morphology and the presence of a long sword process, Rhyacophila aphrodite is a member of the “ Pararhyacophila subgroup C” sensu Waringer et al. 2018 (= “Branch 4- R. vulgaris Group” of Ross 1956; “ R. vulgaris Branch- R. vulgaris Group” of Schmid 1970). Within the framework of the Rhyacophilidae key by Waringer & Graf (2011), R. aphrodite can be separated from all other European Trichoptera species by possessing the following features:
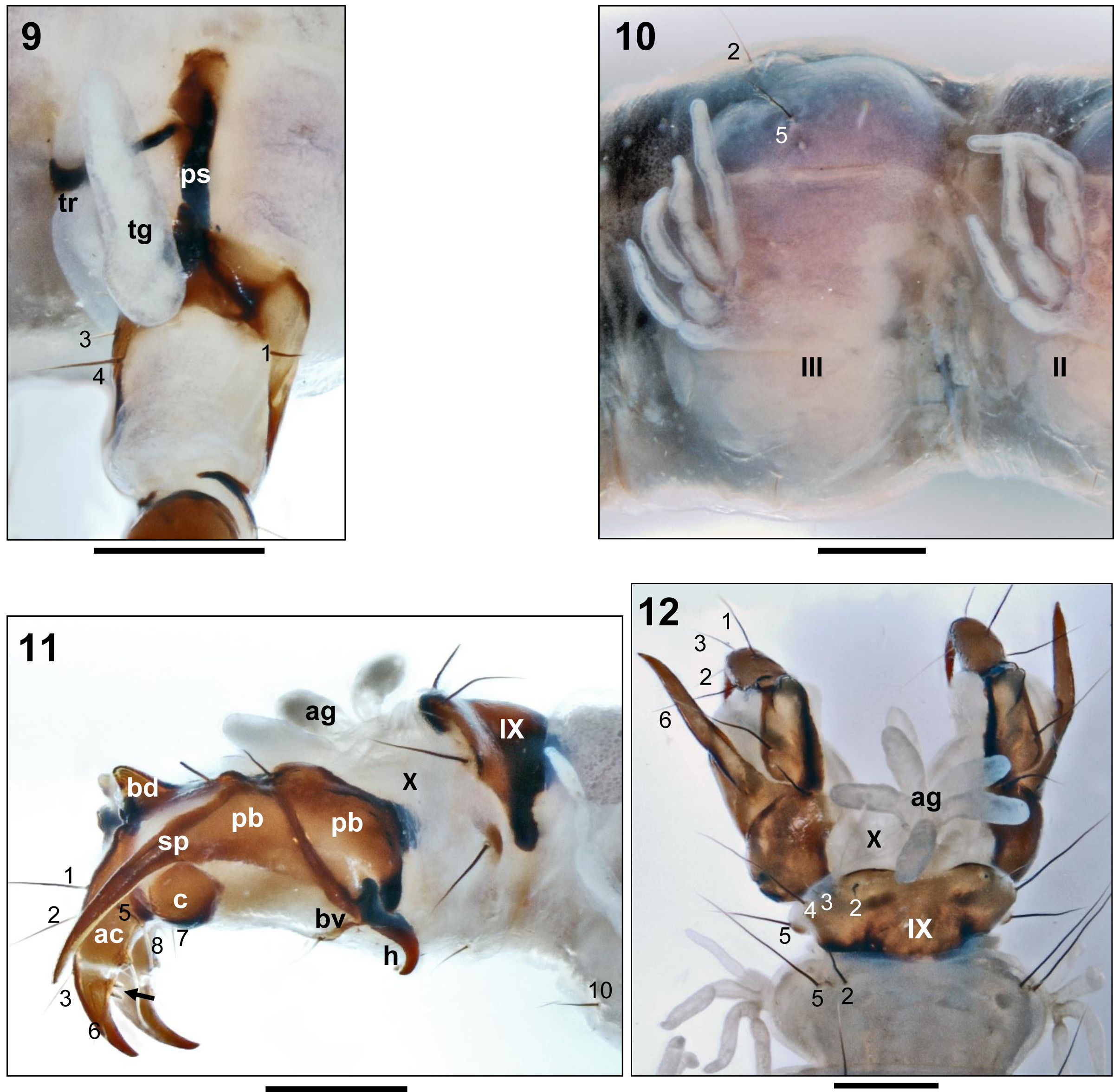
- Only pronotum sclerotised ( Fig. 3 View FIGURES 1–3 pr);
- Labrum sclerotized ( Fig. 1 View FIGURES 1–3 lb);
- Abdominal segment IX with dorsal sclerite ( Figs 11, 12 View FIGURES 9–12 );
- Without transportable case;
- Meso- and metathoraces with monofilament gills ( Fig. 9 View FIGURES 9–12 tg);
- Abdominal segments I to VIII with tetrafilament gills ( Fig. 10 View FIGURES 9–12 );
- Anal prolegs each with sword process more than half as long as anal claw ( Fig. 11 View FIGURES 9–12 sp);
- Distal anal claw tooth perpendicular to anal claw and not bent posterodorsad ( Fig. 11 View FIGURES 9–12 arrow);
- Distal anal claw tooth shorter than half anal claw width at distal tooth insertion ( Fig. 11 View FIGURES 9–12 arrow).
This unique set of characters, combined with the unique status of this species as a micro-endemic of Cyprus, allows a reliable identification of the larvae of R. aphrodite .
Ecology and distribution
Our collection site of Rhyacophila aphrodite was a small, shady stream over volcanic bedrock (Troodos ophiolite from Upper Cretaceous; Geological Survey Department 2019), close to the Byzantine Trooditissa Monastery in the Troodos mountain range between Agios Dimitrios and Pano Platres (Limassol Province). With respect to distribution, R. aphrodite is a micro-endemic species of Cyprus ( Malicky, 2005a, 2005b; Graf et al. 2008; Neu et al. 2018) and one of only 5 endemic species known from this island, highlighting the low degree of endemism of Cypriotic Trichoptera when compared with other islands. The other four endemic species comprise one hydroptilid ( Stactobia urania Malicky 1976b ), one polycentropodid ( Polycentropus milikuri Malicky 1975 ), and two hydropsychids ( Hydropsyche discreta Tjeder 1952 , H. adspersa Navás 1932 ) ( Malicky 2005b). This may be due to the fact that Cyprus is not very isolated and that faunal connections are prominent with Asia Minor, the Caucasus area, and, to some extent, also with Palestine. Interestingly, R. aphrodite belongs to a group of Rhyacophilidae species restricted to Europe and characterized by small, species–specific distribution ranges spread all over the Mediterranean (‘paleomediterranean disjunction type’ sensu Malicky 2005b). It is similar to R. gudrunae Malicky 1972 , a micro-endemic species of Crete, and several west-Mediterranean Rhyacophila taxa in Corsica, Sardinia, the Pyrenees, and the Apennine and Iberian Peninsula.
The streams on Cyprus lack longitudinal zonation patterns observed in other European running waters: the potamal region is lacking due to the restricted stream length, and distinct krenal communities are replaced by an impoverished stream fauna ( Malicky 1976a). However, three island-specific longitudinal zones can be observed on Cyprus: (1) a spring-brook zone with cold water (up to 9°C water temperature) at approximately 1600 m a.s.l. in the Troodos Mountains; (2) an intermediate zone from approximately 1500 down to 900 m a.s.l.; and (3) a zone below 800 m a.s.l. Interestingly, only the intermediate zone (2) has a relatively rich fauna whereas the spring brooks within zone (1) are poor in species, but high in abundance for the few species that occur in them, as demonstrated by Plectrocnemia renetta Malicky 1975. On the other hand, the lowland zones of Cyprus are composed mainly of hydroptilids, in addition to Agapetus caucasicus and Polycentropus milikuri Malicky 1975 (Malicky 1976a). Our larval material of R. aphrodite has been collected in the species-rich intermediate zone (2).
Larval mandible morphology is very close to that of R. rougemonti McLachlan 1880 (Waringer & Malicky 2018), the latter classified as a predator preying on a wide spectrum of aquatic invertebrates (Graf et al. 2008); presumably, R. aphrodite shares this feeding type. With respect to phenology, final instar larvae were present in the streams of Cyprus at the end of April, and the male holotype, together with several adult paratypes, were sampled on 11 May ( Malicky 1975). However, the full duration of the flight period remains unknown; the closely similar species R. gudrunae is on the wing from May to December ( Malicky 2005b).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |