
Salaziella praslinensis Bolotov & Spitsyn
|
publication ID |
https://doi.org/ 10.1080/00222933.2021.1947536 |
|
publication LSID |
lsid:zoobank.org:pub:E3236191-01A7-4DD4-905D-1D236C0F1446 |
|
persistent identifier |
https://treatment.plazi.org/id/03C26C0A-FFDD-FFD9-C425-6791FBB89D4A |
|
treatment provided by |
Plazi |
|
scientific name |
Salaziella praslinensis Bolotov & Spitsyn |
| status |
|
Salaziella praslinensis Bolotov & Spitsyn gen. & sp. nov.
Figures 1–5 View Figure 1 View Figure 2 View Figure 3 View Figure 4 View Figure 5 .
LSID. http://zoobank.org/ urn:lsid:zoobank.org:pub:E3236191-01A7-4DD4-905D-1D236C0F1446
Type material. Holotype female RMBH Sph 0918, SEYCHELLES: Praslin Island, Salazie Trail, 4.3223°S, 55.7324°E, alt. 207 m, mid-altitude palm forest on a hill slope, 16.ii.2016, Bolotov leg. GoogleMaps Paratype female RMBH Sph0919, the same label data as for the holotype GoogleMaps . The type series is deposited in the RMBH – Russian Museum of Biodiversity Hotspots , N . Laverov Federal Center for Integrated Arctic Research of the Ural Branch of the Russian Academy of Sciences , Arkhangelsk, Russia .
Etymology. This species is named after Praslin Island, where it is distributed.
Diagnosis. This species can be distinguished from other Derbidae taxa by having a unique tegmen venation and specific markings pattern (see description below).
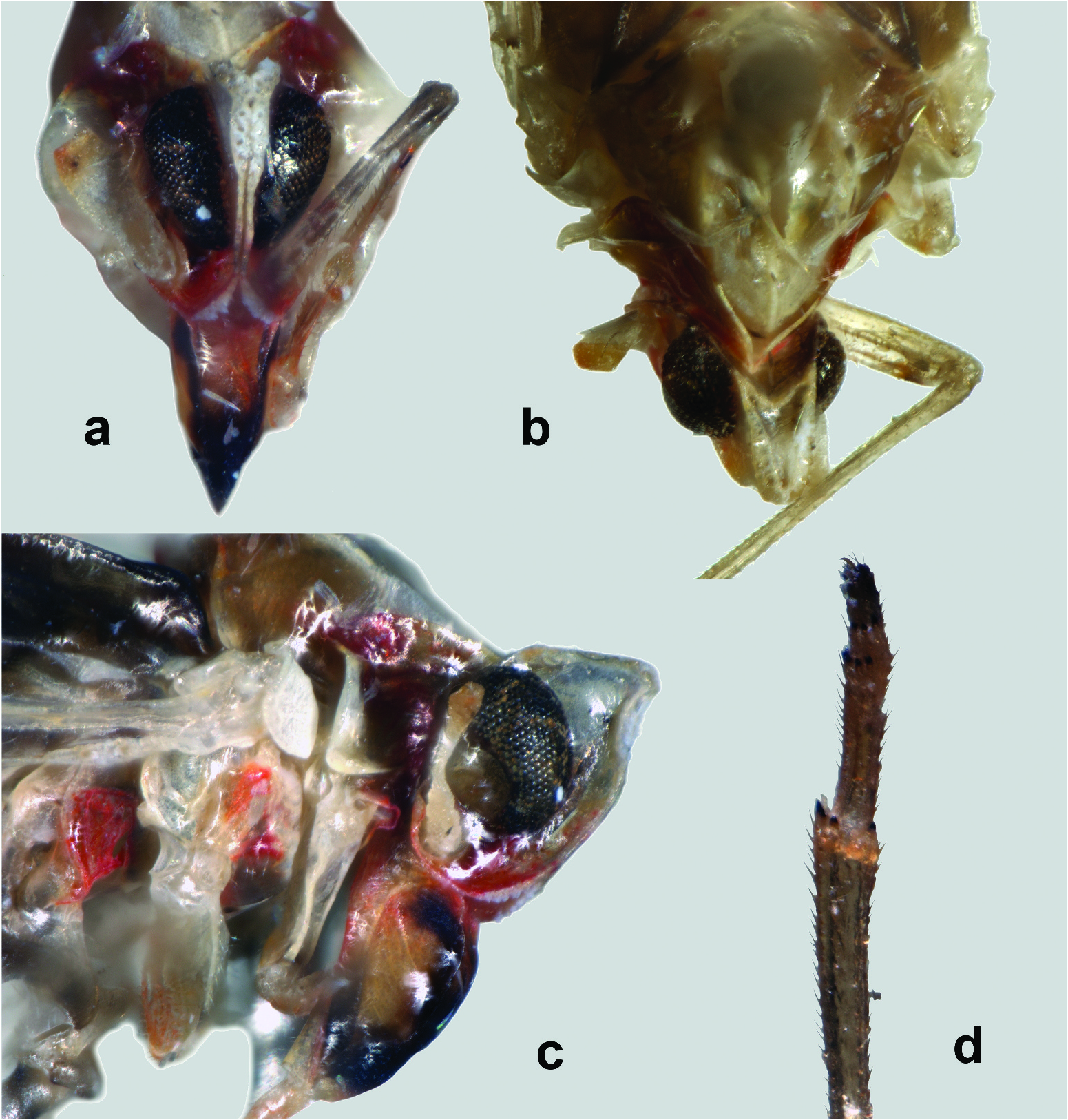
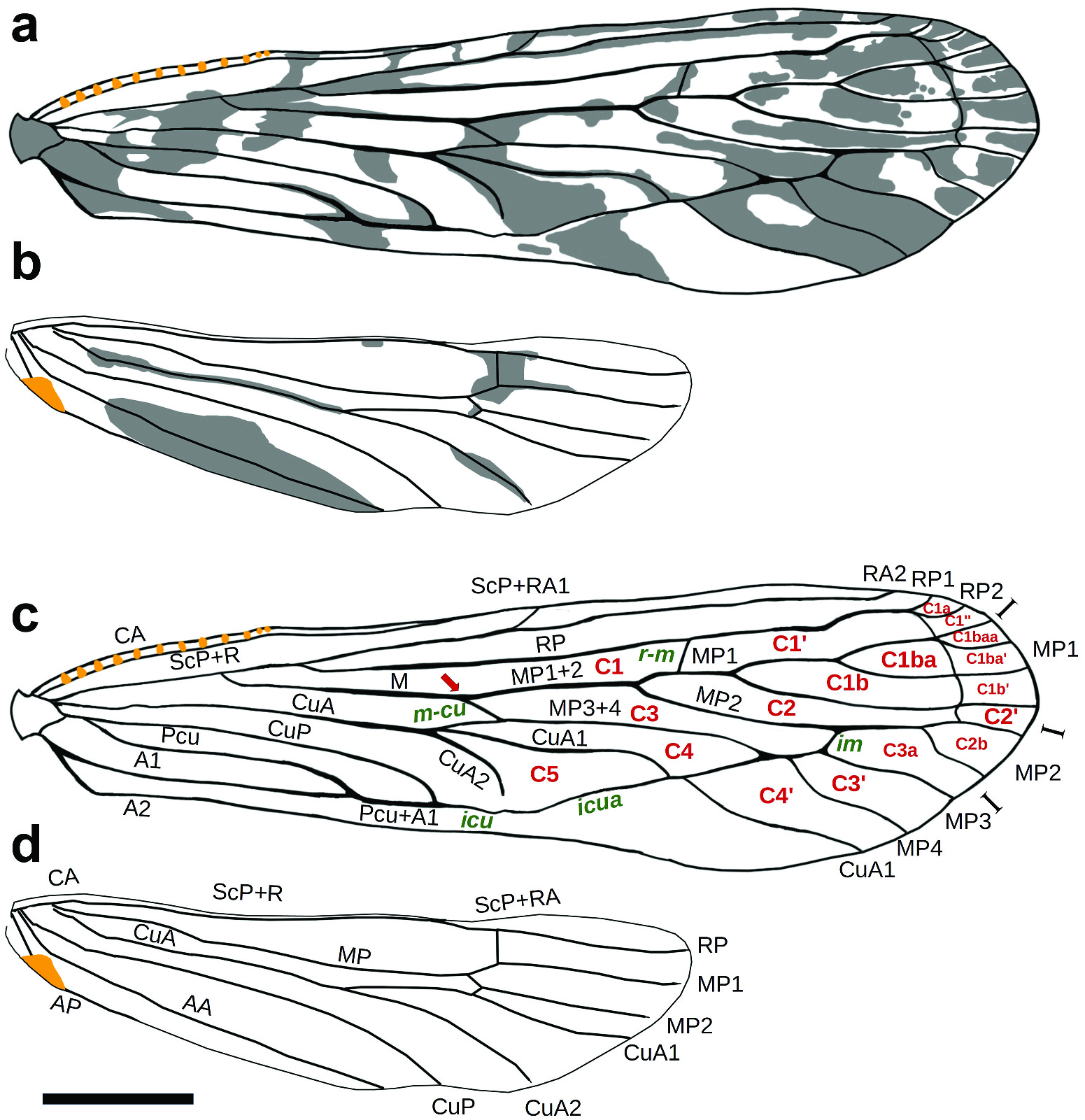
Description. A medium-sized planthopper ( Figure 1 View Figure 1 (a–b)), resting position with wings spread. Body length from tip of head to tip of tegmen 8.4 mm; width of tegmen at level of apex of clavus 1.5 mm; maximum width of tegmen 1.8 mm; length of tegmen from base to tip 7.3 mm (holotype female). Head with eyes much narrower than pronotum, dark red ( Figure 2 View Figure 2 (a–c)). Eyes black, large but not reaching the epistomal suture. Antennae whitish, with reddish apex, slightly shorter than frons, first and second segments long, cylindrical; subantennal process absent. Vertex white, two times as long as broad, tapering anteriorly, with large V-shaped impression dorsally, its lateral margins with rows of pits, anterior margin rounded. Frons white, with gradually widening lateral carinae, forming a broad V-shaped extension in the uppermost part. Clypeus dark red, with black distal end, longer than frons, tricarinate, slightly curved distally, rostrum slightly surpassing the posttrochanters. Pronotum and mesonotum tricarinate. Legs yellowish grey, moderately long. Hind tibia with 4 apical teeth and without lateral spines; 1st and 2nd tarsomeres with 5 apical teeth and without platellae. Body short, black dorsally and ochraceous, with a few reddish stripes laterally. Tegmen ( Figure 3 View Figure 3 (a)): Long, narrow, slightly tapering towards base, with almost straight costal and posterior margins; subhyaline, with a well-developed, diffuse blackish markings pattern. Postcostal cell with four dark stripes; large dark area basally; two short dark bands subbasally; a large dark median spot near the postclaval margin; areola postica almost completely darkened; a large dark areas between CuA1 and MP3; dark spots between veins medially; radial area with numerous unclear dark spots and dark lines along veins. Costal margin, radial and some media posterior (MP2 and MP3) veins tinged with dark red. Tegmen venation ( Figure 3 View Figure 3 (c)): Basal section of costa thick and bears a row of 12 sensory pits. Subcosta posterior + radius vein forked medially into short ScP + RA1 and elongated RA2 without furcation. Radius posterior branch diverged from ScP + R at the basal 1/3 of the tegmen and bifurcated near the apex. Veinlet r-m present. Media posterior vein starts from ScP + R and bifurcated into MP1 + 2 and MP3 + 4; MP1 with four terminals; MP2 with two terminals; MP3 and MP4 without furcation. Cubitus anterior (CuA) arising independently from base of forewing; three branched (CuA and M3 + 4 fused basally via m-cu crossvein); CuA2 not reaching the posterior margin of the tegmen; branches of CuA united for a long distance, forming part of the subapical line of crossveins. Cubital posterior vein not reaching the posterior margin of the tegmen, being fused with CuA branches via icu and icua crossveins. Postcubitus apically fused with A1 (Pcu + A1); A2 forms the claval margin of the tegmen. Clavus rather broad, open (combined CuP + A1 not reaching margin within claval area). Hindwing ( Figure 3 View Figure 3 (b)): Long (approximately 0.65 of the tegmen length), narrow, with straight costal margin and convex posterior margin; subhyaline, with a diffuse blackish markings pattern: a large dark spot around the apex of the medial cell; a broad dark area near the posterior margin; some darkness along CuA vein. Stridulatory plate present, its external margin straight. Hindwing venation ( Figure 3 View Figure 3 (d)): Costa thick. ScP + R vein forked medially into ScP + RA and RP. Media posterior and cubital anterior veins bifurcate. CuP, AA, and AP veins single. Female terminalia and ovipositor ( Figure 4 View Figure 4 (a–g)): Pygofer with a small dorsal lobe and rounded, short dorso-lateral lobes. Gonapophysis VIII broad, rounded apically, with a bund of fine hairs distally. Gonapophysis IX narrow, curved ventrally, slightly pointed apically. Gonoplacs broad, bearing dorsal sclerite with two or more acute teeth. Pregenital sternite bilobate, with U-shaped incision distally. Bursa copulatrix large, ovate, with two thick spines dorsally. Ductus bursae very long, strongly sclerotised, curved ventrally. Male: Unknown.
Distribution. This species is endemic to Praslin, Seychelles. It is unknown beyond the type locality.
Habitat and biology. The habitat of the new species is a patch of mid-altitude palm forest on a hill slope ( Figure 5 View Figure 5 ). The forest is dominated by Palms Phoenicophorium borsigianum (K. Koch) Stuntz , Verschaffeltia splendida H. Wendl. , Nephrosperma vanhoutteanum (H. Wendl. ex Van Houtte) Balf. , and Deckenia nobilis H. Wendl. ex Seem. (Arecaceae) , and Cinnamon Cinnamomum verum J. Presl. (Lauraceae) . All the palms are endemic to the Inner Seychelles ( Matatiken and Dogley 2009). Among them, Phoenicophorium borsigianum is particularly abundant at the type locality. Although host plants of Salaziella praslinensis gen. & sp. nov. are unknown, we could assume that it is almost certainly associated with one or several endemic Seychelles palms. It was shown that most derbids at the adult stage are host specialists on palms ( Attié et al. 2008), e.g. coconut ( Cocos nucifera L.) and oil palm ( Elaeis guineensis Jacq. ) ( Wilson 1987; Yap et al. 2015; Dollet et al. 2020). Phoenicophorium borsigianum is a host plant of Sikaiana albomaculata , the only other native member of the subfamily Otiocerinae from Seychelles ( Löcker et al. 2009). A few additional Seychelles endemic planthoppers are also associated with this palm species, e.g. Aquaelicium typicum Distant, 1917 (Derbidae) , Fipsianus andreae Holzinger, 2009 , and F. picturatus ( Distant, 1917) (Cixiidae) ( Holzinger 2009a, 2009b).
Population abundance. This species seems to be rare, as it was not recorded under earlier comprehensive surveys of the Seychelles Derbidae ( Distant 1917; Löcker et al. 2009). The two type specimens of Salaziella praslinensis gen. & sp. nov. were collected accidentally, when they were attracted by a light source under an awning during heavy tropical rain at the late evening (approx. 8:00 pm). Although one of the authors (I.N.B.) visited the type locality multiple times between February 2016 and January 2020 and searched for additional specimens on palms, these attempts were unsuccessful.
Conservation and threats. We recommend assessing the new species as Critically Endangered (CR B1a), because it is known to occur at only a single palm forest site on
Praslin. The endemic palm forests on Praslin are highly threatened by human activities such as cutting, road construction, and forest fires ( Senterre 2009). Moreover, outbreaks of the invasive Yellow Crazy Ant Anoplolepis gracilipes Smith, 1857 on Praslin during the last decade could threaten Salaziella praslinensis gen. & sp. nov. Currently, this ant species heavily affects animal communities in the Vallée de Mai National Park, another endemic palm forest site on Praslin ( Kaiser-Bunbury et al. 2014). Anoplolepis gracilipes was found to be abundant in the type locality of the new species (Ivan Bolotov, pers. observ., 2016–2020).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |