
Schizopera abei, Karanovic & Kim & Grygier, 2015
|
publication ID |
https://doi.org/ 10.1080/00222933.2015.1028112 |
|
DOI |
https://doi.org/10.5281/zenodo.4324079 |
|
persistent identifier |
https://treatment.plazi.org/id/70424C4F-FF8D-0D44-01C9-FCD063FBFD5D |
|
treatment provided by |
Carolina |
|
scientific name |
Schizopera abei |
| status |
sp. nov. |
Schizopera abei sp. nov.
( Figures 1–7 View Figure 1 View Figure 2 View Figure 3 View Figure 4 View Figure 5 View Figure 6 View Figure 7 )
Type locality
Japan, Shiga prefecture, Nagahama city, Nomura-cho township, Ane River , a few meters upstream from the older of the two Nomurabashi bridges, cobble and large pebble shoal, interstitial water from Karaman– Chappuis hole, 35°24’56.3”N 136°19’26.8”E GoogleMaps .
Specimens examined
Holotype female ( LBM1430005760 View Materials ) dissected on one slide, allotype male ( LBM1430005761 View Materials ) dissected on one slide, four paratype females ( LBM1430005762 View Materials - LBM1430005765 View Materials ) and three paratype males ( LBM1430005766 View Materials - LBM1430005768 View Materials ) dissected on one slide each, 36 paratypes (13 males, nine females, and 14 copepodids) ( LBM1430005769 View Materials ) together in ethanol, and 21 paratypes (10 males and 11 females; LBM1430005773 View Materials ) together on one SEM stub; all collected from type locality, 28 May 2012, leg. M.J. Grygier, S. Kanao, and S. Nakaoka (sample no. 71–01).
Six paratypes (one male and five females) together in ethanol ( LBM1430005770 View Materials ) from Japan, Shiga prefecture, Hikone city, Inukata township, Inukami River , fine sand in river bed, interstitial water pumped with Bou-Rouch style pump, 35°14’5.9”N 136°15’1.9”E, 26 May 2011, leg. K. Tanida party (sample no. 17–02) GoogleMaps .
One paratype male in ethanol ( LBM1430005771 View Materials ) from Japan, Shiga prefecture, Hikone city, Obori township, Seri River , fine sand in river bed, interstitial water pumped with Bou-Rouch style pump, 35°14’48.0”N 136°16’5.4”E, 25 May 2011, leg. K. Tanida party (sample no. 2–08) GoogleMaps .
Two paratype females together in ethanol ( LBM1430005772 View Materials ) from Japan, Shiga prefecture, Taga town, Nakagawara, Seri River , fine sand in river bed, interstitial water pumped with Bou-Rouch style pump, 35°14’9.9”N 136°16’56.9”E, 25 May 2011, leg. K. Tanida party (sample no. 19–01) GoogleMaps .
Etymology
The new species is named in honour of Dr. Yuji Abe, curator at the Taga Town Museum, Japan, in recognition of his continuing help in collecting subterranean copepods in and around Taga. The name is a noun in the genitive singular.
Description
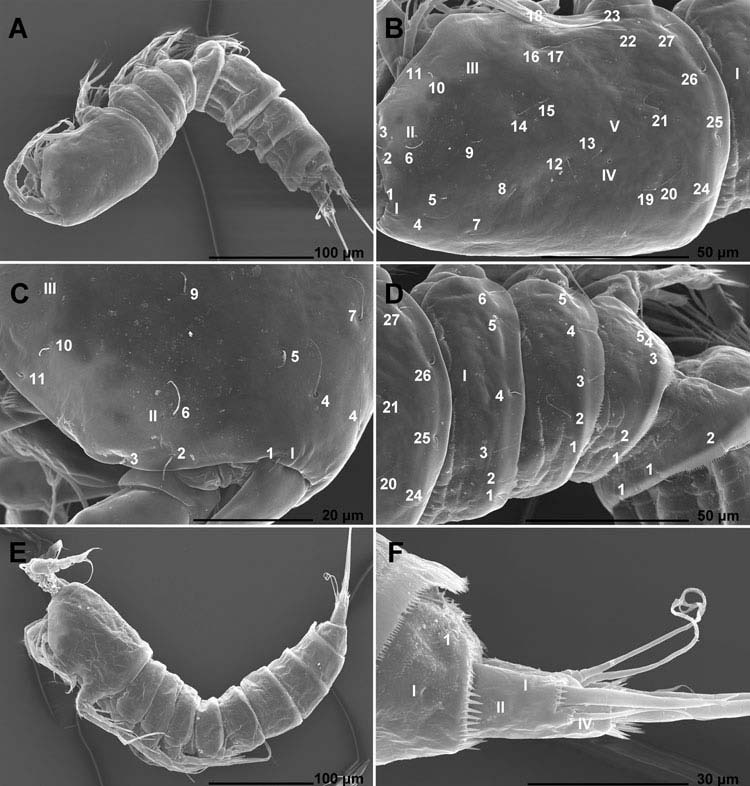
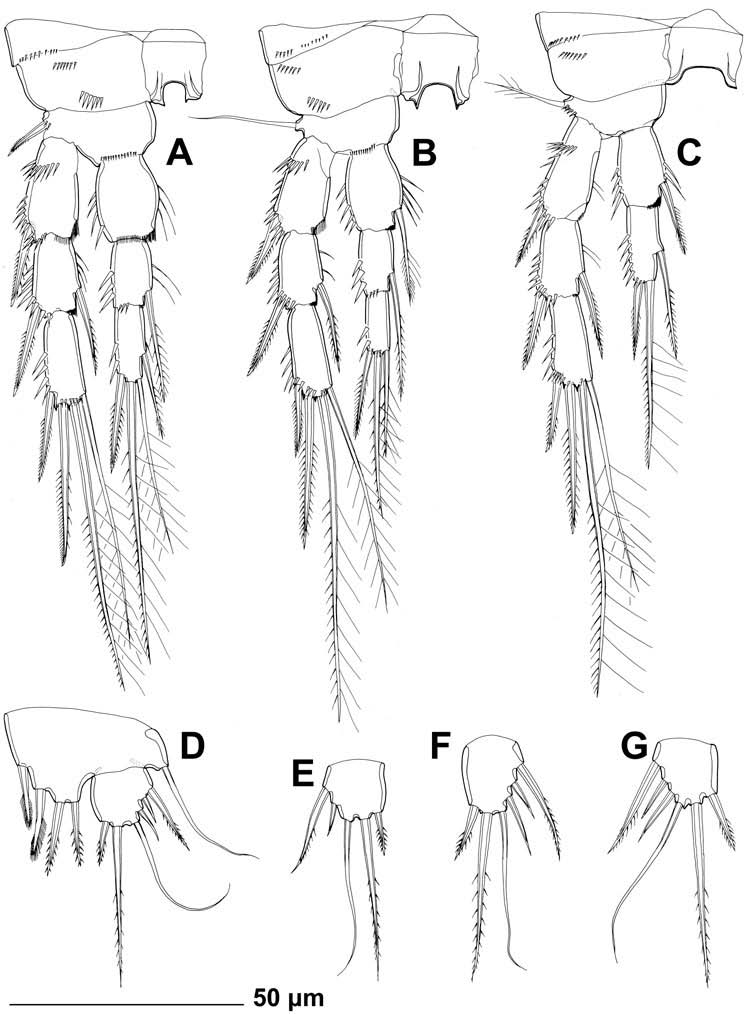
Female (data from holotype and 16 paratypes). Total body length, measured from tip of rostrum to posterior margin of caudal rami (excluding caudal setae and appendages), from 330 to 355 μm. Colour of preserved specimens yellowish. Nauplius eye not visible. Prosome comprising cephalothorax with completely fused first pedigerous somite and three free pedigerous somites; urosome comprising fifth pedigerous somite, genital double-somite (fused genital and first abdominal somites) and three free abdominal somites. Habitus ( Figure 1A View Figure 1 ) cylindrical but not particularly slender, without distinct demarcation between prosome and urosome; prosome/urosome ratio about 1.1 (in dorsal view); greatest width at posterior end of cephalothorax but difficult to establish, with cephalothorax only slightly wider than genital doublesomite. Body length/width ratio about 4.5. Free pedigerous somites without pronounced lateral or dorsal expansions. Integument of all somites relatively well sclerotised, generally very smooth, without cuticular windows or pits. All somites (except cephalothorax) and caudal rami, besides other ornamentation, with two or more parallel rows of minute spinules. Hyaline fringe of all somites broad and at least partly serrated, except for smooth posterior end of cephalothoracic shield. Surface of somites, rostrum and caudal rami with total of 70 pairs of cuticular organs (15 pairs of cuticular pores and 55 pairs of sensilla; see Figures 1–4 View Figure 1 View Figure 2 View Figure 3 View Figure 4 ).
Rostrum ( Figures 1C View Figure 1 , 5A View Figure 5 ) long and clearly demarcated at base, reaching two thirds of second antennular segment, linguiform, about twice as long as wide, with two short spiniform processes on tip and single dorsal pair of sensilla (R-1) at about midlength.
Cephalothorax ( Figure 1A–C View Figure 1 ) about 1.3 times as long as wide in dorsal view (without rostrum); representing 30% of total body length, tapering towards anterior end in dorsal view only in anterior third. Hyaline fringe of cephalothoracic shield wide and smooth. Cephalothoracic shield ( Figure 1B, C View Figure 1 ) with five pairs of pores (C-I to C-V) and 27 pairs of sensilla (C-1 to C-27); sensilla C-16 and C-17 very close to each other, as well as sensilla C-19 and C-20; pores C-I very close to sensilla C-1 (both at base of rostrum) and pores C-II relatively close to sensilla C-6; pores C-IV more than twice as large as any other prosomal pores; sensilla C-19 to C-27 and pores C-XI and C-XII probably belonging to the first pedigerous somite, incorporated into cephalothorax; lateral marginal zone includes sensilla C-2, C-3, C-11, C-18, and C-23 (see Figure 1B View Figure 1 ); posterior marginal zone including sensilla C-24 to C-27 (see Figure 1D View Figure 1 ).
Pleuron of first free prosomite (second pedigerous somite; Figure 1D View Figure 1 ) with two rows of minute spinules in anterior half, one pair of anterior dorsal-lateral pores (FP1-I) and six pairs of long sensilla (FP1-1 to FP1-6); posterior marginal zone including sensilla FP1-1, FP1-2 and FP1-4 to FP1-6, while sensilla pair FP1-3 situated slightly more anteriorly and longer than any other pair; sensilla FP1-1 and FP1-6 probably serially homologous to sensilla C-24 and C-27, respectively, on first pedigerous somite; other serial homologies difficult to determine; hyaline fringe wide, serrated only dorsally, smooth dorso-laterally and laterally.
Pleuron of second free prosomite (third pedigerous somite; Figure 1D View Figure 1 ) slightly longer and with several more short rows of minute spinules than pleuron of first free prosomite, without pores, with only five pairs of long sensilla (FP2-1 to FP2-5); posterior marginal zone including sensilla FP2-1 and FP2-3 to FP2-5, while sensilla pair FP2-2 situated slightly more anteriorly and longer than any other pair; dorsalmost pair of sensilla more widely spaced than on pleuron of first free prosomite but recognition of serially homologous pairs relatively easy (FP2-1 = FP1-1, FP2-2 = FP1-3, FP2-3 = FP1-4, FP2-4 = FP1-5 and FP2-5 = FP1-5); hyaline fringe wide, serrated dorsally and dorso-laterally, smooth laterally.
Pleuron of third free prosomite (fourth pedigerous somite; Figure 1D View Figure 1 ) slightly narrower and significantly shorter (especially dorsally) than pleuron of second free prosomite, but with similar number of minute spinules and also without pores, with only five pairs of long sensilla (FP3-1 to FP3-5); all sensilla probably serially homologous to their counterparts with same Arabic numerals on pleuron of second free prosomite; hyaline fringe narrow dorsally, wide laterally, serrated dorsally and dorsolaterally, smooth laterally.
First urosomite (fifth pedigerous somite; Figure 1D View Figure 1 ) about as long as pleuron of third free prosomite but with fewer minute spinules, with one pair of ventro-lateral pores near base of fifth legs (U1-I) and two pairs of dorsal sensilla (U1-1 and U1-2); sensilla pair U1-1 probably serially homologous to sensilla pair FP3-1, but serial homology of sensilla pair U1-2 not obvious (perhaps FP3-3?); hyaline fringe wide and finely serrated dorsally and dorso-laterally, smooth only near base of fifth legs.
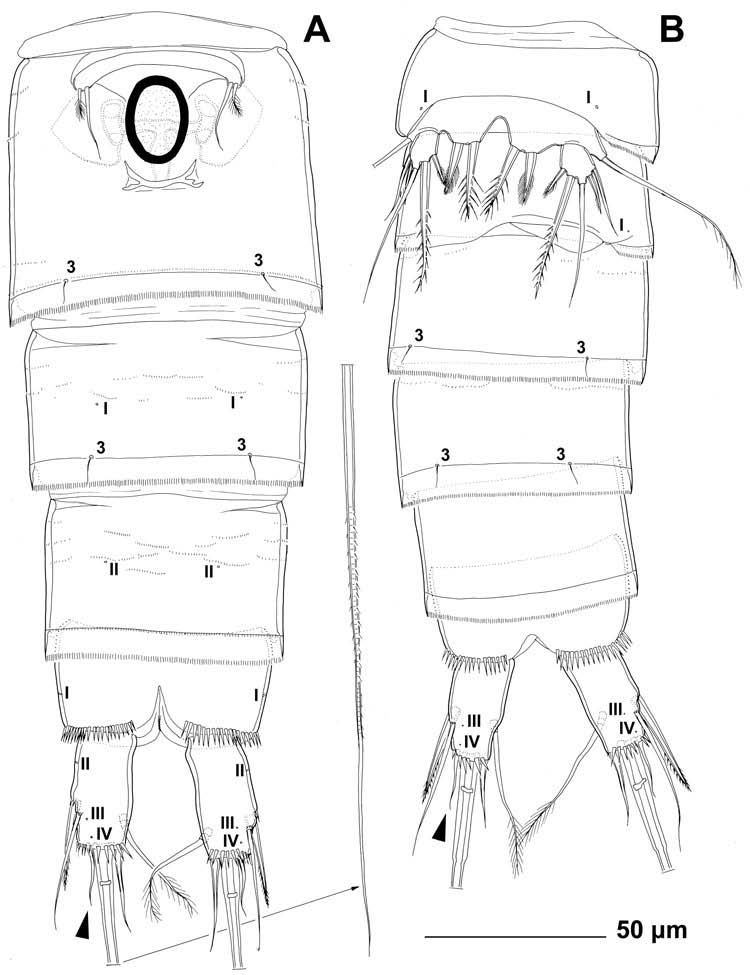
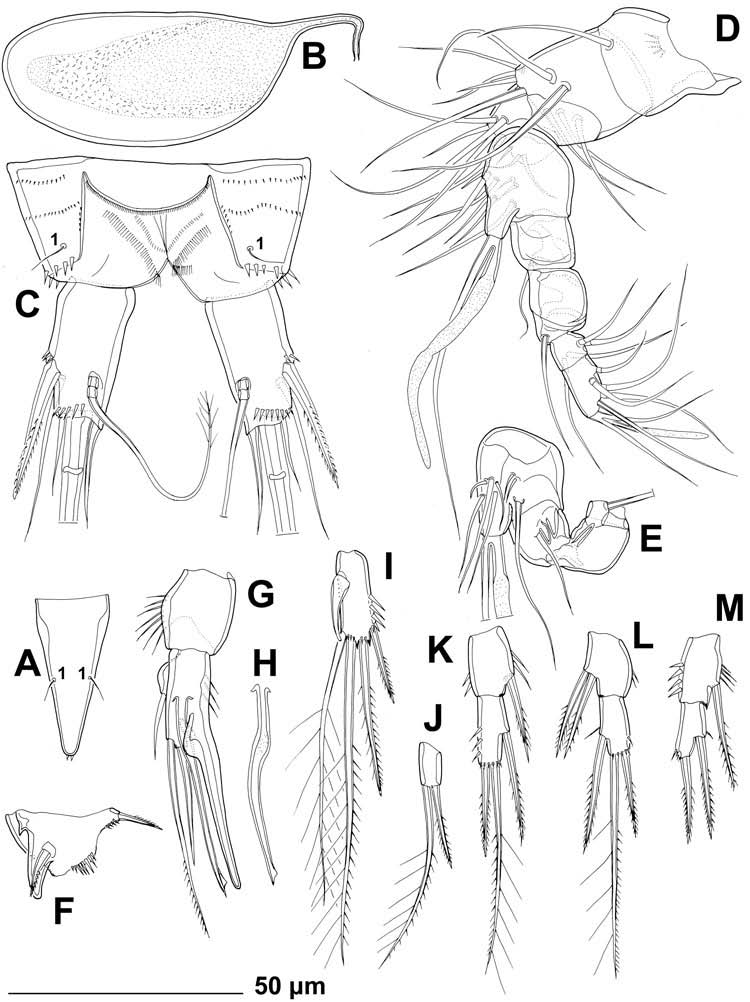
Second urosomite ( Figures 1A View Figure 1 and 4A View Figure 4 ) in female almost completely fused with third urosomite into genital double-somite, with three dorsal and dorso-lateral parallel rows of minute spinules and only two pairs of dorsal posterior sensilla (U2-1 and U2-2), both probably serially homologous to their counterparts with same Arabic numerals on first urosomite. No remnants of hyaline fringe. Female genital complex ( Figure 4A View Figure 4 ) with single copulatory pore posterior to epicopulatory bulb (latter also serving as copulatory duct), two small seminal receptacles placed inside large, paired, genital apertures; apertures with two ventral gonopores, each covered by reduced sixth leg. Epicopulatory bulb large, ovoid, strongly sclerotised, about 1.4 times as long as wide. Seminal receptacles very small, kidney-shaped, not reaching anterior margin of epicopulatory bulb, about 0.6 times as long as epicopulatory bulb.
Third urosomite ( Figures 1A View Figure 1 and 4A View Figure 4 ) with posterior row of minute spinules, wide and finely serrated hyaline fringe, and three pairs of posterior sensilla: one dorsal (U3-1), one lateral (U3-2) and one ventral (U3-3); establishing serially homologous sensilla of third and second urosomites not easy (possibly, U3-1 = U2-2). Genital double somite as a whole about 0.8 times as long as wide (ventral view), with only small internal ridge dorso-laterally (but no external suture) indicating original segmentation.
Fourth urosomite ( Figures 1A View Figure 1 and 4A View Figure 4 ) slightly narrower and significantly shorter than genital double-somite, with several anterior rows of minute spinules, wide and finely serrated hyaline fringe, one pair of anterior ventral pores (U4-I), and three pairs of posterior sensilla (U4-1 to U4-3); all sensilla with homologous pairs on third urosomite (i.e., U4-1 = U3-1, U4-2 = U3-2, and U4-3 = U3-3) but ventral pair (U4-3) slightly closer together.
Fifth urosomite (preanal; Figures 1A View Figure 1 and 4A View Figure 4 ) narrower than third urosomite but not shorter, without sensilla, with one pair of posterior dorsal pores (U5-I), one pair of anterior ventral pores (U5-II), several short and slightly arched rows of minute spinules in anterior half, and posterior continuous row of minutes spinules; ventral pores serially homologous with those on fourth urosomite (i.e., U5-II = U4-I); hyaline fringe sharply serrated, ventrally as wide as that in third urosomite but dorsally extended into wide and long pesudoperculum, this nearly reaching posterior margin of anal somite, with about 35 sharp teeth.
Sixth urosomite (anal; Figures 1A View Figure 1 and 4A View Figure 4 ) slightly narrower and only about 0.7 times as long as fifth urosomite, cleft medially in posterior half, with one pair of large dorsal sensilla (U6-1), one pair of large lateral pores (U6-I), posterior row of large spinules at base of each caudal ramus, and several short curved rows of minute spinules (mostly on dorsal and lateral surfaces); anal operculum short, narrow, convex, situated anterior to dorsal sensilla, completely covered by pseudoperculum, with posterior row of numerous hair-like minute spinules, representing 55% of somite’ s width; anal sinus widely opened, without any chitinous projections, with weakly sclerotised walls and two diagonal rows of long, hair-like spinules.
Caudal rami ( Figures 1A View Figure 1 and 4A View Figure 4 ) strongly sclerotised, about 1.9 times as long as greatest width in ventral view, almost cylindrical (somewhat tapering towards caudal end in posterior third but with almost straight inner margin), with space between them slightly less than one ramus width; ornamented with posterior row of large spinules, several smaller spinules at base of lateral setae, and four pairs of pores: anterior dorso-lateral pores (CR-I), anterior ventro-lateral pores (CR-II) and two pairs of posterior ventral pores (CR-III and CR-IV); armed with six elements (two lateral, one dorsal and three apical). Dorsal seta slender and apically pinnate, about 1.3 times as long as ramus, inserted at about 2/3 of ramus length in deep recess, triarticulate at base (i.e. inserted on two pseudojoints). Lateral proximal spine stout, bipinnate, inserted at 2/3 of ramus length and 0.8 times as long as ramus. Lateral distal seta slender, smooth, inserted slightly ventrolaterally at 3/4 of ramus length, and about as long as ramus. Inner apical seta with wide and strong base, smooth, about 0.7 times as long as ramus. Inner principal apical seta with breaking plane, very strong, distally pinnate, about six times as long as caudal ramus. Outer principal apical seta smooth, slender and very short, only about half as long as caudal ramus (arrowed in Figure 4A View Figure 4 ).
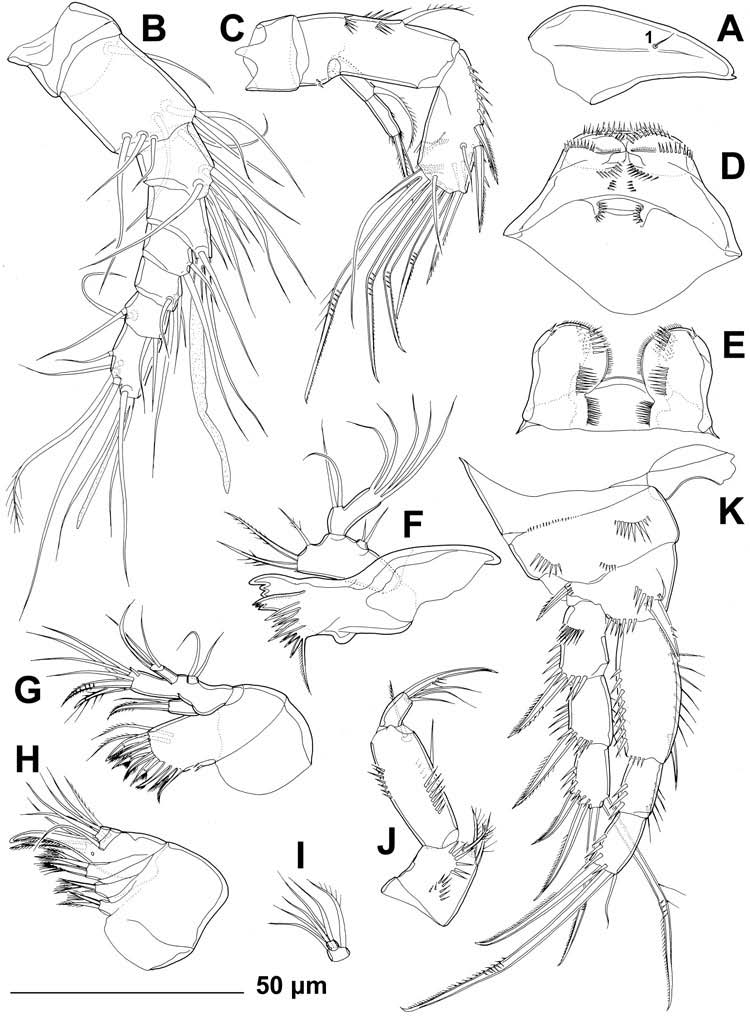
Antennula ( Figure 5B View Figure 5 ) eight-segmented, approximately half as long as cephalothorax, with slender aesthetasc on eighth segment fused basally to two apical setae, large aesthetasc on fourth segment reaching significantly beyond tip of appendage and fused basally to slightly shorter seta, and setal formula 1.9.8.3.2.4.4.7. Two lateral setae on seventh segment and four on eighth segment biarticulate (i.e. inserted on short pseudojoint). All setae slender, all except for one on eighth segment smooth, and most ending apically in pore (except apical and subapical setae); apical pores only observable under scanning electron microscope. Length ratio of antennular segments, from proximal end and along caudal margin, 1:2.2:0.8:0.9:0.8:0.8:0.8:1.6.
Antenna ( Figure 5C View Figure 5 ) comprising coxa, basis, two-segmented endopod and much smaller but also two-segmented exopod. Coxa short, 0.7 times as long as wide, without ornamentation or armature. Basis and first endopodal segment partly fused along posterior surface. Basis also short and unarmed, about 0.6 times as long as wide, ornamented with small spinules along inner margin. First endopodal segment twice as long as wide and 2.8 times as long as basis, with two arched rows of large spinules in proximal half on outer margin, and one short, unipinnate lateral seta at middle. Second endopodal segment 1.2 times as long as first, more slender proximally, with two surface frills distally; lateral armature consisting of two strong spines flanking small, slender seta; apical armature consisting of seven elements: one smooth, slender, short seta, one unipinnate short spine and four geniculate setae, longest fused basally to another smooth and slender but long seta; all geniculate setae with minute spinules along outer (concave) margin distally, longest one with several long spinules along inner (convex) margin as well. Ornamentation of second endopodal segment consisting of longitudinal row of large spinules along anterior margin and diagonal row of large spinules between lateral and apical armature elements. Both exopodal segments of about same width and length; first segment armed with 1 unipinnate subapical seta, unornamented; second segment ornamented with transverse apical row of slender spinules and several small lateral spinules, armed apically with one smooth, slender seta and one strong, bipinnate spine of about same length as former, both about 1.5 times as long as segment.
Labrum ( Figure 5D View Figure 5 ) large, trapezoidal, rigidly sclerotised, with slightly concave cutting edge, ornamented with numerous slender apical and subapical spinules, as well as several rows of spinules of various length and orientation along posterior surface.
Paragnaths ( Figure 5E View Figure 5 ) slightly smaller than labrum, also rigidly sclerotised, almost linguiform, connected by medial trapezoidal lobe resembling labrum in shape, with numerous spinules along inner and apical margins (apical ones much more robust), as well as two longitudinal rows of spinules on anterior surface.
Mandibula ( Figure 5F View Figure 5 ) with cutting edge of coxa narrow, armed with two complex teeth in ventral part (both tricuspidate), eight simple (unicuspidate) teeth in dorsal part, and one unipinnate dorsalmost seta; coxa unornamented. Basis smaller and shorter than coxa, about twice as long as wide, armed with three bipinnate slender setae along inner margin; ornamented with several minute spinules at base of ventralmost seta. Endopod one-segmented, twice as long as wide, armed with two lateral and five apical smooth setae. Exopod very small but distinct segment, armed with two smooth apical setae.
Maxillula ( Figure 5G View Figure 5 ) with large praecoxa, arthrite highly mobile, armed apically with six strong, unipinnate spines, and two dorsalmost unipinnate setae; armed laterally with two smooth, slender setae and ornamented with short row of spinules at base of arthrite. Coxa small, armed with two setae on inner margin; distal seta slender and smooth, proximal seta very strong, spiniform and bipinnate. Inner margin of basis furnished with two strong, curved, bipinnate spines and five smooth, slender setae; distalmost seta minute, others as long as spine or longer. Endopod onesegmented, small, about twice as long as wide, armed with three apical smooth setae, innermost seta longest and strongest. Exopod also distinct but very small segment, half as long as wide, armed with two slender and smooth apical setae.
Maxilla ( Figure 5H, I View Figure 5 ) composed of syncoxa, basis and two-segmented endopod. Syncoxa unornamented, large, ovoid, with three endites, proximal and central ones each armed with two subequal setae, distal one armed with two setae and one spine, all pinnate near distal tip. Basis much smaller than coxa, elongate, armed with one apical claw-like spine (partly fused to basis), one unipinnate and strong apical seta and one smooth, slender lateral seta on anterior surface; ornamented with one pore on posterior surface. Endopod very small, short and wide, armed with three slender setae on each segment.
Maxilliped ( Figure 5J View Figure 5 ) prehensile, three-segmented, composed of coxobasis and two-segmented endopod. Coxobasis 1.2 times as long as wide, cylindrical, ornamented with two arched rows of large spinules on anterior margin, armed with three strong, unipinnate setae on inner (median) margin, all about half as long as coxobasis. First endopodal segment about 2.5 times as long as wide and 1.6 times as long as coxobasis, ornamented with two longitudinal rows of large spinules on anterior surface and one row of smaller spinules on posterior surface; armed with two short, slender setae, one centrally on inner margin and other subapically on posterior surface. Second endopodal segment smallest, only 0.3 times as long as first and twice as long as wide, armed apically with one claw-like unipinnate spine and three smooth, slender setae; spine more than twice as long as second endopodal segment and 1.3 times as long as longest seta.
All swimming legs ( Figures 1A View Figure 1 , 5K View Figure 5 and 6A–C View Figure 6 ) slender, short in comparison to body length and width, composed of small unarmed triangular praecoxa, large unarmed quadrate coxa, smaller armed basis, three-segmented armed exopod, and three-segmented armed endopod. Coxae in all pairs of legs connected by unornamented intercoxal sclerite. All exopodal and endopodal segments of about same length, except for much longer first endopodal segment of first leg.
First swimming leg ( Figure 5K View Figure 5 ) with small, short and wide intercoxal sclerite, concave at distal end and unornamented. Praecoxa ornamented with posterior row of minute spinules on anterior surface. Coxa also ornamented with several short arched rows of spinules of various sizes on anterior surface. Basis with one inner and one outer strong, pinnate spine, inner one stronger and about 1.5 times as long as outer; ornamentation consisting of several spinules at base of each spine and one additional row of large spinules along distal margin between endopod and exopod, all on anterior surface. Exopod armed with single outer-distal spine on first and second segments, and with two outer spines and two apical geniculate setae on third segment; all exopodal segments ornamented with strong spinules along outer margin and subdistally, and additionally along inner margin of second segment; first exopodal segment with additional arched row of strong spinules on anterior surface proximally; inner geniculate seta on third segment slightly longer than entire exopod and about 1.3 times as long as outer geniculate seta. Endopod geniculate, with first segment 0.65 times as long as entire exopod, 2.4 times as long as second endopodal segment, about twice as long as wide; third endopodal segment about 1.1 times as long as second endopodal segment; endopodal armature consisting of one strong but short inner seta on first segment (inserted at about 4/5), and three setae on third segment [innermost slender and smooth, middle longest and geniculate, outermost spiniform seta (or spine?) 0.6 times as long as middle one]; endopodal ornamentation consisting of strong spinules along outer margin of all segments, and also along inner margins of first two segments.
Second swimming leg ( Figure 6A View Figure 6 ) with even smaller praecoxa than in first leg, also ornamented with posterior row of spinules on anterior surface. Coxa ornamented with two short horizontal rows of large spinules on anterior surface. Intercoxal sclerite with paired, pointed, distal protrusions. Basis armed only with outer bipinnate spine, ornamented with small spinules at base of outer spine and with minute spinules along distal margin at base of endopod. Distal inner corners of first and second exopodal and endopodal segments with serrated hyaline frills. All exopodal and endopodal segments ornamented with strong spinules on outer margins; first and second segments also with weaker spinules along inner margins. Exopod armed with outer-distal spine on first and second segments, inner spiniform seta on second segment, two outer spines and two apical setae on third segment; all spines and setae strong and bipinnate; outer apical seta on third segment appearing transitional in form between spine and seta, with outer margin furnished with short spinules and inner margin with long, slender spinules. Endopod slightly shorter than exopod, armed with single inner seta on second segment, and four elements on third segment: outer-distal short spine, two apical long setae and one inner strong seta (inserted at 2/3).
Third swimming leg ( Figure 6B View Figure 6 ) very similar to second, except basis armed with outer slender seta instead of spine, first endopodal segment with one inner seta, and third endopodal segment with only three elements (inner seta missing).
Fourth swimming leg ( Figure 6C View Figure 6 ) similar to third leg, except endopod twosegmented and only about 0.6 times as long as exopod, innermost armature element on ultimate endopodal segment representing inner seta (inner distal seta absent), and intercoxal sclerite without pointed processes.
Fifth leg ( Figure 6D–G View Figure 6 ) unornamented, biramous, composed of large, broad baseoendopod and small, ovoid exopod, with division line visible on anterior surface but not complete on posterior surface. Baseoendopod with outer basal smooth seta arising from relatively short setophore. Endopodal lobe almost triangular, extending to middle of exopod, armed with four very stout, spiniform elements (two inner ones probably spines, two outer ones probably spiniform setae); length ratio of endopodal armature elements, from inner side, 1:1.1:1.2:1. Exopod from 0.9 to 1.25 times as long as maximum width, normally armed with six elements: two innermost apical ones strong and bipinnate, outer apical one smooth and slender, distal and central outer ones short, smooth and spiniform, and proximal outer one long, strong and pinnate; length ratio of exopodal armature elements, from inner side, 1:3:3:1:0.8:1.8. One exopod with only five elements ( Figure 6E View Figure 6 ).
Sixth leg ( Figure 4A View Figure 4 ) indistinct, very small cuticular plate covering gonopore, armed with two slender setae; inner seta smooth, about 2.4 times as long as outer bipinnate seta.
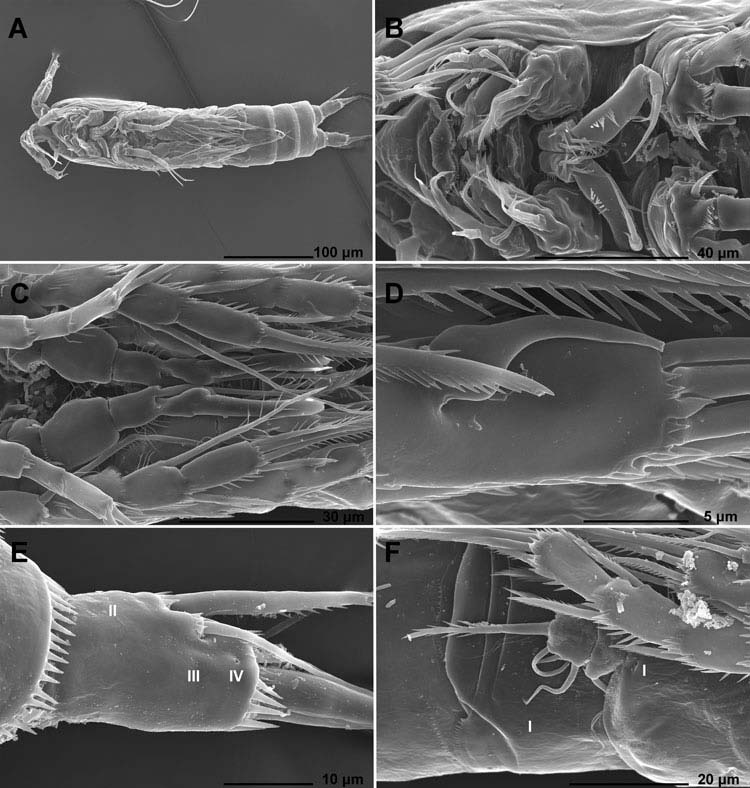
Male (data from allotype and 14 other paratypes). Body length ranging from 324 to 347 μm. Habitus ( Figures 1E View Figure 1 , 2A View Figure 2 and 3A View Figure 3 ) slightly more slender than in female, but also cylindrical, and with similar proportions of prosome/urosome, and cephalothorax/genital somite. Body length/width ratio about 4.2. Ornamentation of rostrum ( Figure 7A View Figure 7 ), prosomites ( Figure 2B–D View Figure 2 ) and first urosomite ( Figures 2D View Figure 2 , 3F View Figure 3 and 4B View Figure 4 ), as well as colour and nauplius eye, as in female.
Genital somite ( Figures 2E View Figure 2 , 3F View Figure 3 and 4B View Figure 4 ) more than twice as wide as long. Single, completely formed, longitudinally placed spermatophore ( Figure 7B View Figure 7 ) inside first two urosomites in most specimens. Ornamentation consists of two pairs of large dorsal sensilla as in female (U2-1 and U2-2) and additional pair of ventral pores at base of sixth legs (U2-I).
Third urosomite ( Figures 2F View Figure 2 , 4B View Figure 4 ) ornamented as in female, but not fused to second (genital) urosomite.
Fourth and fifth urosomites ( Figures 2E, F View Figure 2 and 4B View Figure 4 ) as in female, except ventral pair of pores absent and fewer minute spinules present.
Sixth urosomite ( Figures 1F View Figure 1 , 2F View Figure 2 , 3E View Figure 3 , 4B View Figure 4 and 7C View Figure 7 ) as in female.
Caudal rami ( Figures 1F View Figure 1 , 2F View Figure 2 , 3E View Figure 3 , 4B View Figure 4 and 7C View Figure 7 ) slightly shorter in comparison with anal somite and more slender than in female but without any difference in armature or ornamentation. Outer principal seta also very short and slender (arrowed in Figure 4B View Figure 4 ).
Antennula ( Figure 7D, E View Figure 7 ) half as long as cephalothorax, strongly prehensile and nine-segmented (basically, female’ s sixth segment subdivided in male), with geniculations between fourth and fifth and seventh and eighth segments. Segments participating in geniculations strengthened with cuticular plates along anterior surface, with largest such plates on fifth segment. Aesthetascs as in female, on fourth and last segments; that on fourth segment somewhat wider than in female. First two and last two segments similar to female. Setal formula: 1.9.8.9.1.0.1.4.7. Most setae smooth and with pore on tip; same setae biarticulate as in female.
Antenna, labrum ( Figure 3B View Figure 3 ), mandibula ( Figure 3B View Figure 3 ), maxillula ( Figure 3B View Figure 3 ), maxilla ( Figure 3B View Figure 3 ), maxilliped ( Figure 3B View Figure 3 ), exopod and endopod of first swimming leg ( Figure 3C View Figure 3 ), exopod of second swimming leg ( Figure 3C View Figure 3 ), endopod of third swimming leg and fourth swimming leg ( Figures 3F View Figure 3 and 7K View Figure 7 ) as in female.
First swimming leg ( Figures 3B View Figure 3 and 7F View Figure 7 ) with modified basis, inner margin very rigidly sclerotised, with spiniform smooth distal process and smaller sharp process at its base. Inner spine on basis smaller than in female, without spinules at its base, inserted more proximally and about as long as larger spiniform process.
Second swimming leg ( Figures 3C View Figure 3 and 7G, H View Figure 7 ) with transformed endopodal second and third segments. Second segment with part of inner margin protruded as rounded indistinct lobe, without ornamentation on its surface; inner seta shorter than in female, smooth and slender. Third segment completely modified; inner apical seta unipinnate and longer than in female, outer apical seta smooth and strong, outer apical spine transformed into smooth, lanceolate implement with slightly swollen part at about first third of its length and abruptly tapering tip; outer distal corner produced into long, blunt spiniform process, about as long as lanceolate outer spine. As result of these transformations, third segment medially cleft.
Third swimming leg ( Figures 3D View Figure 3 and 7I View Figure 7 ) with very characteristic element on anterior surface of third exopodal segment probably representing hugely enlarged tubular pore: swollen in basal part, with pore on tip, inserted at 2/5 and close to inner margin, reaching distal margin of third segment.
Fifth legs ( Figures 3F View Figure 3 and 4B View Figure 4 ) with medially fused baseoendopods. Endopodal lobe much smaller and shorter, trapezoidal, extending to first third of exopod in length, armed with two very strong apical spines; inner spine about 1.5 times as long as outer one and with fewer but stronger spinules. Exopod about as long as wide, demarcated basally on both anterior and posterior surfaces, armed with only five elements (one short lateral element missing); length ratio of exopodal armature elements, from inner side, 1:3.8:4:1.1:2.1.
Sixth legs ( Figures 3F View Figure 3 and 4B View Figure 4 ) expressed as pair of small, short cuticular plates, without armature or ornamentation; left one better demarcated at base and probably functioning as genital flap.
Variability
In addition to the slight variability in body length (see above), several other features were observed as variable. The exopod of the female fifth leg was about as long as wide in most specimens (see Figure 6G View Figure 6 ), but the length/width index could vary from 0.9 to 1.25 ( Figure 6D, F View Figure 6 ); one paratype female was observed with only five elements on one fifth leg exopod ( Figure 6E View Figure 6 ), while the opposite leg has a normal condition of six elements ( Figure 6D View Figure 6 ). One paratype male had only two elements on the third endopodal segment of the third leg ( Figure 7J View Figure 7 ), while the opposite leg and all other specimens examined had the normal condition of three elements. Another paratype male had an abnormal endopod of the fourth leg, with two inner setae on the first segment ( Figure 7L View Figure 7 ), while its opposite leg is normal (i.e. same as in the female; Figure 7K View Figure 7 ). One paratype male had only two elements on the second endopodal segment of the fourth leg ( Figure 7M View Figure 7 ), while its opposite leg was normal.
Molecular results
DNA was extracted, and the mtCOI fragment successfully PCR-amplified from three specimens of our new species ( Table 1). All sequences were translated into protein using MEGA and were shown to have no evidence of stop codons, ambiguities or insertions/deletions indicative of non-functional copies of mtCOI. BLAST analyses of GenBank revealed that the obtained sequences are copepod in origin and not contaminants. All analyses were run with all additional 37 Schizopera mtCOI sequences downloaded from GenBank, and with four sequences belonging to two outgroup species ( Table 1). The complete mitochondrial genome of S. knabei (see Easton et al. 2014) was trimmed for the largest overlapping range with any other congener after alignment (640 bases), while all other sequences were used with their original lengths, ranging from 407 to 639 bases. Our alignment showed no gaps. Average pairwise distances between species were found to be very high ( Table 2), with the lowest divergence (16.9%) between S. leptafurca and S. uranusi . Divergences between all other taxa were in excess of 18%. These high divergence values are generally indicative of distinct species by comparison with other crustaceans ( Lefébure et al. 2006) and other harpacticoid copepods ( Karanovic and Cooper 2011a, 2012; Karanovic and Kim 2014; Karanovic et al. 2014). There was evidence for multiple divergent lineages (12.2% average sequence divergence; 4.9–17% divergence between haplotypes) within the species S. akation , but the question remains as to whether these lineages represent the presence of cryptic species or are just divergent mtDNA sequences within a species. All other divergences within morphospecies were below 2%, and these are indicative of intraspecific variability ( Lefébure et al. 2006). Our three sequences of S. abei did not show any differences, which is not surprising considering that all three specimens were collected in the same locality and are probably kin.
The ingroup was recovered in all three cladistics analyses ( Figure 8), and was supported with relatively high bootstrap values: 90% for ML, 83% for MP and 99% for NJ. All our analyses also supported the presence of at least 13 genetically divergent ingroup lineages, corresponding to 13 morphospecies, and all seven of the multi-sample lineages were supported with high bootstrap values (between 90 and 100%). A sister group relationship between S. uranusi and S. leptafurca was only relatively weakly supported, despite the overwhelming morphological evidence suggesting this (see Karanovic and Cooper 2012), but the monophyly of a group comprising S. uranusi , S. leptafurca and S. emphysema was moderately well supported, in accordance with morphological evidence. A relatively well-supported sister group relationship between S. analspinulosa s. str. and S. analspinulosa linel (95% in our NJ analysis) was also shown. Although morphological evidence suggests a relatively close relationship among S. kronosi , S. analspinulosa s. str. and S. analspinulosa linel , this relationship was only recovered in our NJ analysis, and with extremely low bootstrap support (24%). Most basal ingroup clades had very low bootstrap support, which is likely to be the result of the low phylogenetic resolution of the mtCOI gene in basal nodes of the tree, possibly due to saturation at third codon positions ( Karanovic and Cooper 2012) and also to various lengths of the fragments amplified (see Table 2). For example, our MP analysis resulted in a single most parsimonious tree of 494 steps, but the consistency and retention indices were low (0.41 and 0.77 respectively), as was the number of parsimony-informative sites (270), while the number of variable sites was 298 and there were 28 singletons. Not surprisingly, the tree topology differed between different methods employed. For example, our NJ analysis suggested S. sp. 2 as a sister clade to all other Schizopera , while our MP analysis grouped this species together with S. abei and S. knabei (albeit with a bootstrap support of only 27%), and our ML analysis suggested an unresolved basal position ( Figure 8). The one specimen that did not match our morphospecies (JQ390571.1; preliminary identification as S. cf. uranusi ) formed a separate lineage and is likely to represent an uncharacterised species of Schizopera .
The phylogenetic position of S. abei among its other congeners could not be established based on the cladistic analysis of mtCOI with any confidence. Our ML analysis suggested a sister relationship with the American marine S. knabei , but this clade was very weakly supported (30%) and was not recovered in our NJ and MP analyses. Instead, our MP analysis suggested a sister relationship with S. sp. 2 (13% bootstrap support) with them as a sister clade to S. knabei , while our NJ analysis suggested S. abei as a sister clade to a large group of nine species (25% bootstrap support). This all suggests that our new Japanese species has no close relatives among the congeners for which we had mtCOI sequences. This is not surprising as most of them come from inland waters of Australia. None of our analyses suggested a particularly close relationship of S. abei and S. akolos , the only two species in this group with a two-segmented endopod of the fourth leg, that would have been considered members of the genus Schizoperopsis based on morphological characters proposed by Apostolov (1982).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
SubPhylum |
Crustacea |
|
Class |
|
|
SubClass |
Copepoda |
|
Order |
|
|
Family |
|
|
Genus |