
Smeringopina pulchra (Millot, 1941)
|
publication ID |
https://doi.org/10.11646/zootaxa.3713.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:C5F0BC11-92C0-4B30-9DB3-200882AC8950 |
|
DOI |
https://doi.org/10.5281/zenodo.6161965 |
|
persistent identifier |
https://treatment.plazi.org/id/B20287ED-FFED-FF91-B990-C5AEFDC13DED |
|
treatment provided by |
Plazi |
|
scientific name |
Smeringopina pulchra (Millot, 1941) |
| status |
|
Smeringopina pulchra (Millot, 1941) View in CoL
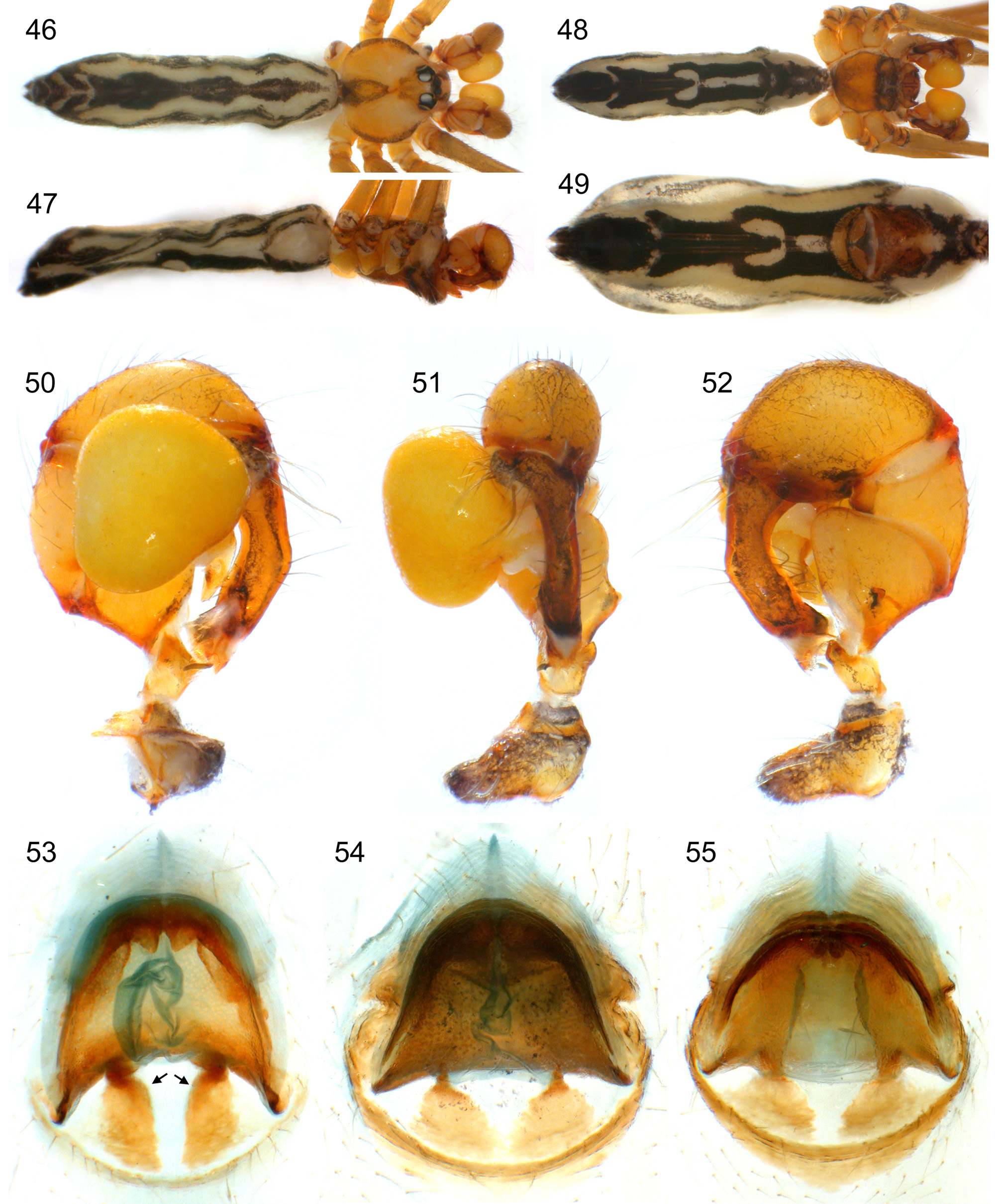
Figs. 5 View FIGURES 2 – 16 , 34–40 View FIGURES 34 – 45 , 53 View FIGURES 46 – 55 , 56–75 View FIGURES 56 – 62 View FIGURES 63 – 75
Smeringopus pulcher Millot 1941: 20 –22, figs. 8A–K.
Smeringopina pulchra: Huber 1995: 299 (transfer to Smeringopina ). Huber & Fleckenstein 2008: fig. 14 (SEM of comb hairs).
Types. ♂ holotype from Ivory Coast, Bingerville [ 5°21.6’N, 3°54’W], ix.1937 (J. Millot), in MNHN (Ar 10482), with original label: “ Sm. pulcher M., Type, J. Millot rec. et. det., Bingerville, C. d’Iv., IX 37 ”. 6♂ 6♀ 2 juvs. paratypes, same data as holotype, in MNHN (Ar 10476, 10477, 10479). 1♂ 2♀ 2 juvs. paratypes from Ivory Coast, Man [ 7°24.6’N, 7°33’W], ix.1937 (J. Millot), in MNHN (Ar 10478, 10493). 1♂ paratype from Guinea, Macenta [ 8°32.5’N, 9°28.2’W], viii.1937 (J. Millot), in MNHN (Ar 10481); all types examined.
Notes. Millot (1941) did not explicitly designate a holotype in his publication but he separated a male and labeled it as “ Type ”. This is here considered the holotype.
The male paratype from Macenta ( Guinea) is somewhat dubious (labeling error?) because S. pulchra could not be found in any Guinean locality during a recent expedition (only S. guineensis was found in eastern Guinea).
Other material examined. IVORY COAST: Lamto [ 6°13.2’N, 5°01.8’W], 17./ 28.vi.1963, collector not given, 1♂ 5♀ (2 vials) in MNHN. Abidjan [ 5°22.2’N, 4°03.0’W], 17.vi.1958 (E.S. Ross, R.E. Leech), 1♂ in CAS. Adiopo Doumé, Centre Suisse de Recherche Scientifique (CSRS), small dry forest, beating, 25.ii.2010 (D. van den Spiegel, R. Jocqué), 1♂ 7♀ 1 juv. in MRAC ( 230358 part); same locality, 25./ 26.ii.2010, sieving litter, 2♂ 3♀ + juvs. (2 vials) in MRAC ( 230222, 230360 part). Appouesso, Forêt classée de la Bossematié [ 6°36.9’N, 3°27.1’W], dense forest, in webs between tree buttresses, xi.1993 (R. Jocqué), 2♂ in MRAC (177658); same data but 30.xi.1993, 1♂ in MRAC (177613 part); same locality, 20.xi.1993, dense forest, transect, 1♂ 1♀ in MRAC (177664); same locality, dense forest, 19.xi.1993, 1♂ 1♀ in MRAC (177651); same locality, rain forest, in web of Euprosthenops [ Pisauridae ], 18.xi.1994, 2♀ in MRAC ( 201073); same locality, rain forest, 24.ii.1997 (R. Jocqué, L. Baert), 1♀ in MRAC (205414 part). Bettié, Forêt classée de Mabi [ 6°04.2’N, 3°24.6’W], dense forest, by hand, 23.xi.1993 (R. Jocqué), 1♀ in MRAC (177635); same locality, forest road, on car, 25.xi.1993 (R. Jocqué), 1♂ 1♀ in MRAC (177696). Forêt des Marais-Tanoé, Aboisso, Dohouan [ 5°10’N, 2°50’W], beating, 25.x.2010 (A. Kablan), 1♀ in MRAC (233748). Guiglo [ 6°32.4’N, 7°28.8’W], viii.1979 (Mission R.U. Gent), 1♀ in MRAC (152038). Forêt de Taï, station du Centre de Recherche Ecologique (CRE), forest across river [ 5°50’N, 7°21’W], near pitfalls III, forest on clayey soil, beating, 20.ii.2010 (M. Diarassouba, R. Jocqué), 1♂ 1♀ in MRAC ( 230287); same locality, forest near camp, by hand, 21.ii.2010 (R. Jocqué), 1♀ in MRAC ( 230136); same locality, road near Chimpanzee Camp, beating in forest, 22.ii.2010, 2 juvs. in MRAC ( 230346 part); same locality, 20.ii.2010, 1♀ 1 juv. in MRAC ( 230470); same locality, at foot of trees, by hand, 22.ii.2010 (R. Jocqué), 1♂ in MRAC ( 230146 part); same locality, forest E of camp, layon Gérard, inundated forest, sieved litter, 20.ii.2010 (R. Jocqué, L. Oulaï), 1♂ 3♀ in MRAC ( 230243).
GHANA: Western Region: Ankasa National Park ( 5°13.0’N, 2°39.1’W), 180 m a.s.l., forest near entrance, day collecting, 22.ii.2013 (B.A. Huber), 10♂ 8♀ in ZFMK (Ar 10163); same data but night collecting, 6♂ 3♀ in ZFMK (Ar 10164); same data, 3♀ in pure ethanol, in ZFMK (Gha 158); Ankasa National Park (~ 5°15.1’N, 2°38.4’W), ~ 100 m a.s.l., forest along Big Tree Trail, 23.ii.2013 (B.A. Huber), 12♂ 6♀ in ZFMK (Ar 10165). Central Region: Kakum National Park ( 5°20.9’N, 1°23.0’W), 160 m a.s.l., forest near entrance, day collecting, 19.–20.ii.2013 (B.A. Huber), 32♂ 13♀ (3 vials) in ZFMK (Ar 10166-68); same data but night collecting, 20.ii.2013 (B.A. Huber), 5♂ 1♀ in ZFMK (Ar 10169); same data, 2♀ in pure ethanol, in ZFMK (Gha 148). Kakum Forest [ 5°21.4’N, 1°23.4’W], primary forest, 16./21./23./ 25.xi.2005 (R. Jocqué, D. de Bakker, L. Baert), 1♂ 7♀ (4 vials) in MRAC (217711, 726, 735, 741); same data but secondary forest, 15./19./ 25.xi.2005, 1♂ 2♀ 1 juv. (3 vials) in MRAC (217172, 704, 731); same data but beating between primary and secondary forest, 11.xi.2005, 1♀ 1 juv. in MRAC (217275). Eastern Region: Atewa Hills, Atewa Atwirebu Reserve ( 6°13.8’N, 0°33.5’W), 740 m a.s.l., 25.ii.2013 (B.A. Huber), 15♂ 9♀ in ZFMK (Ar 10170); same data, 2♀ in pure ethanol, in ZFMK (Gha 134). Atewa Atwirebu Reserve ( 6°13.8’N, 0°32.4’W), 500 m a.s.l., degraded forest along road, 24.ii.2013 (B.A. Huber), 4♂ 6♀ in ZFMK (Ar 10171). Brong Ahafo Region: Booyem ( 7°40.6’N, 1°57.1’W), 470 m a.s.l., at Bibri waterfall, 4.iii.2013 (B.A. Huber), 3♂ 4♀ in ZFMK (Ar 10172); same data, 2♀ in pure ethanol, in ZFMK (Gha 129). Booyem ( 7°39.9’N, 1°57.4’W), 450 m a.s.l., at large rocks, 4.iii.2013 (B.A. Huber), 1♀ in ZFMK (Ar 10173). Volta Region: Agumatsa Wildlife Sanctuary, Wli waterfall ( 7°06.2’N, 0°36.0’E), ~ 300 m a.s.l., forest near waterfall, 27.ii.2013 (B.A. Huber), 1♂ 9♀ in ZFMK (Ar 10174). Tagbo waterfall ( 7°00.7’N, 0°34.4’E), ~ 500 m a.s.l., forest near waterfall, 28.ii.2013 (B.A. Huber), 6♂ 3♀ in ZFMK (Ar 10175).
TOGO: Missahohe [ 6°56’N, 0°36’E], 6.viii.1969 (F. Puylaert), 1♂ in MRAC (136079); same locality, 1893 (E. Baumann), 2♂ in ZMB.
Diagnosis. Distinguished from similar congeners ( S. guineensis , S. bineti ) by straight procursus ( Figs. 40 View FIGURES 34 – 45 , 57 View FIGURES 56 – 62 ), obtuse bulbal apophysis ( Fig. 58 View FIGURES 56 – 62 ), absence of median apophyses proximally on male chelicerae ( Figs. 59 View FIGURES 56 – 62 , 64 View FIGURES 63 – 75 ), and by strongly curved posterior margin of anterior epigynal plate ( Figs. 37 View FIGURES 34 – 45 , 60 View FIGURES 56 – 62 , 73 View FIGURES 63 – 75 ); from S. guineensis also by presence of ventro-distal apophysis on procursus ( Fig. 57 View FIGURES 56 – 62 ), all modified hairs on large cheliceral apophyses grouped close together ( Figs. 59 View FIGURES 56 – 62 , 68 View FIGURES 63 – 75 ), and by pair of humps on anterior epigynal plate ( Fig. 66 View FIGURES 63 – 75 ); from S. bineti also by longer legs and by absence of dark lateral marks on carapace (only dark margins; Fig. 34 View FIGURES 34 – 45 ).
Male (Appouesso, MRAC 177664). Total body length 6.8, carapace width 1.5. Leg 1: 68.3 (15.3 + 0.7 + 15.3 + 33.9 + 3.1), tibia 2 missing, tibia 3: 7.3, tibia 4: 9.7; tibia 1 L/d: 115. Distance PME-PME 170 µm, diameter PME 160 µm, distance PME-ALE 80 µm, distance AME-AME 35 µm, diameter AME 135 µm. Carapace ochre-orange with brown triangular mark posteriorly and brown lateral margins; ocular area posteriorly brown, clypeus with indistinct pair of brown lines and brown rim, sternum light ochre-brown with darker margins; legs ochre, tips of femora and tibiae whitish, dark rings subdistally on femora and tibiae and in patella area; abdomen ochre-gray with distinct black pattern dorsally, laterally, and ventrally. Habitus as in Figs. 34–36 View FIGURES 34 – 45 , ocular area slightly elevated, secondary eyes with indistinct ‘pseudo-lenses’ ( Fig. 67 View FIGURES 63 – 75 ); clypeus unmodified except evenly curved sclerotized rim; deep thoracic pit and pair of shallow furrows diverging behind pit ( Fig. 65 View FIGURES 63 – 75 ). Chelicerae as in Fig. 59 View FIGURES 56 – 62 , without median projections proximally, with lateral proximal apophyses, large distal apophyses provided with 3–5 modified hairs each, without distal apophyses close to fang joint. Palps as in Figs. 38–40 View FIGURES 34 – 45 ; coxa only slightly bulged retrolaterally; trochanter with simple retrolatero-ventral apophysis; femur with retrolateral projection, with large ventral bulge distally, without ventral or prolateral modification proximally; prolateral femur-patella joint only slightly moved toward ventrally; tarsus with about 3–4 stronger hairs; procursus without hinge, straight, with two long ventral hairs bent around procursus and directed toward dorsally, distally with distinctive ventral apophysis and moveable dorsal sclerite embedded in membranous cuticle ( Figs. 56–57 View FIGURES 56 – 62 , 70 View FIGURES 63 – 75 ); bulb with simple obtuse apophysis arising from membranous basal projection ( Fig. 58 View FIGURES 56 – 62 ; sperm duct apparently opens at membranous basal projection at basis of apophysis; Fig. 69 View FIGURES 63 – 75 ). Legs without spines and curved hairs, with few vertical hairs, retrolateral trichobothrium on tibia 1 at 1.5%; prolateral trichobothrium present on all tibiae; pseudosegments barely visible. ALS with seven spigots each ( Fig. 71 View FIGURES 63 – 75 ); gonopore with two epiandrous spigots ( Fig. 72 View FIGURES 63 – 75 ).
Variation. Number of modified hairs on male chelicerae slightly variable; color of sternum variable (ochreorange to dark brown). Tibia 1 in 30 other males: 10.1–16.4 (mean 14.2). The type specimens are bleached but mostly in fair condition; the holotype lacks the right palp and the moveable sclerite distally on the procursus is missing in the left palp (probably broken); the male paratype from Man lacks the right palp; the male paratype from Macenta is in poor condition, with palps and chelicerae detached and partly destroyed.
Female. In general similar to male; sternum and clypeus variably dark. Tibia 1 in 30 females: 9.5–11.5 (mean 10.6). Epigynum consisting of large anterior plate with deep posterior indentation and pair of low humps in anterior part, and large posterior plate composed of sclerotized posterior arc and pair of distinct anterior plates ( Figs. 37 View FIGURES 34 – 45 , 53 View FIGURES 46 – 55 , 60 View FIGURES 56 – 62 , 66, 73 View FIGURES 63 – 75 ); internal genitalia as in Fig. 62 View FIGURES 56 – 62 , with distinct globular structures between uterus externus and anterior epigynal plate ( Fig. 61 View FIGURES 56 – 62 ). ALS as in male ( Fig. 74 View FIGURES 63 – 75 ).
Natural history. In southern Ghana, S. pulchra was often among the dominant web-building spiders, both in well-preserved forests and in degraded forest fragments. In Ankasa N.P. I counted about 60 adult specimens among the buttresses of a single Piptadeniastrum africanum tree. The domed webs were found mainly in protected places but often also in low vegetation. Small nematocerous flies were often found hanging from the webs, sometimes in high numbers. In some places (most notably Ankasa N.P.) Argyrodes spiders were also found in the webs. In Kakum N.P., the spiders were observed to stick strongly to their own silk (when caught with the hand); this was not the case at other localities. When disturbed, the spiders vibrated with high amplitude and then tried to escape, but did not drop to the ground.
Distribution. Widely distributed in southern Ivory Coast, Ghana, and Togo ( Fig. 33 View FIGURE 33 ). The single record from Guinea is dubious (see Notes above).





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |