
Tanapseudes sinensis Bamber, 2000
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3142.1.1 |
|
persistent identifier |
https://treatment.plazi.org/id/AF5F87C3-DEF0-FF68-B19F-C680FCA6941C |
|
treatment provided by |
Felipe |
|
scientific name |
Tanapseudes sinensis Bamber, 2000 |
| status |
|
Tanapseudes sinensis Bamber, 2000 View in CoL
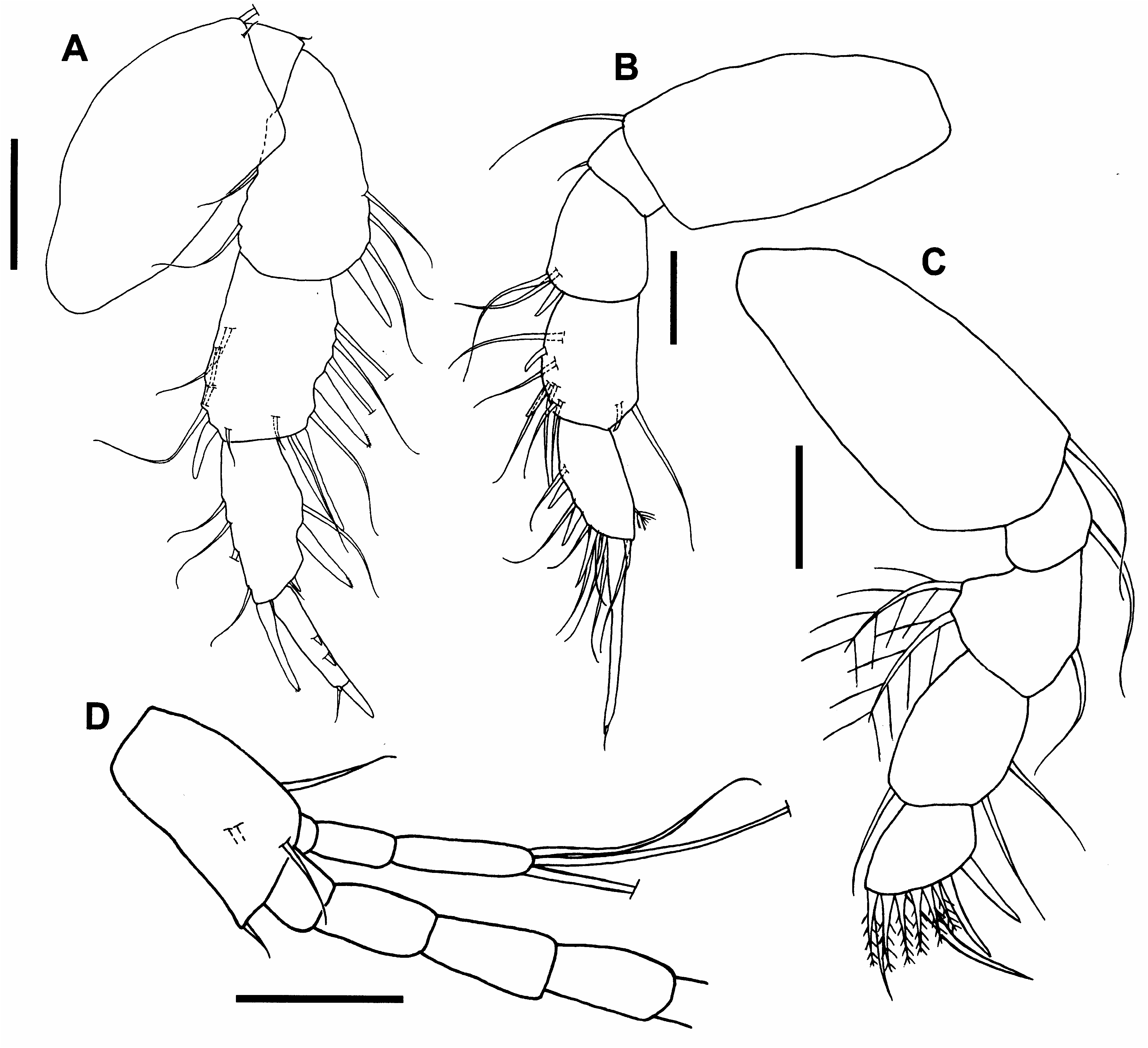
(Figs 109–110)
Tanapseudes sinensis Bamber, 2000: 45 View in CoL , Figs. 4–6. Guţu and Angsupanich, 2005: 51, 52, 54.
Material examined. Allotype: adult female, NHM 1998.2551, Tai Tam Bay , Hong Kong, 22°13'N, 114°14'E, 30m, coll. R.G. Ongche, det. R. Bamber, July, 1993. GoogleMaps
Diagnosis. Pleonites with less than five lateral plumose setae. Pleotelson rounded posteriorly. Male pereopod
1 with long dorsodistal spiniform seta on propodus. Male cheliped carpus lacking ventrodistal rounded protuberance. Pereopods 2–5 with ventral margins of merus and carpus not heavily setose. Uropod basal article with inner distal spiniform projection.
Type locality. South China Sea ( Fig. 100 View FIGURE 100 ) .
Remarks. Hansknecht et al. (2002) synonymized T. sinensis with T. ormuzana , but Guţu and Angsupanich (2005) revalidated it. This is the largest species of Tanapseudes (> 3 mm) and can be distinguished from the other two congeners by the uropod basal article having a spiniform projection on the inner distal corner ( Fig. 110D View FIGURE 110 ).
Ecology. Little is known of the ecology of most members of Kalliapseudidae . The feeding behavior of two species [ P. granulosus (subfamily Tanapseudinae ) and M. macsweenyi (subfamily Kalliapseudinae )], which distinctly differ in mouthpart morphology, was described by Drumm (2005). Based on the observations of Drumm (2005), M. macsweenyi constructs “tubes” in soft sediments using mucus secretions and feeds by filtering detritus and diatoms with plumose setae attached to the chelipeds and maxillipeds. In contrast P. granulosus , which lacks a permanent domicile, appears to be fossorial and feeds by scraping the organic material (e.g. microflora) off sand particles (Drumm, 2005).
A vast majority of the species within the suborder Apseudomorpha appear to be fossorial (e.g., Apseudidae , Sphyrapidae ) or epibenthic (e.g., some Pagurapseudidae and Metapseudidae ). In contrast, some members of the families Kalliapseudidae (Kalliapseudinae) , Parapseudidae Guţu, 1981 , and possibly the small and poorly known Numbakullidae Guţu and Heard, 2002 appear to occupy permanent or semipermanent tubes or burrows. Members of the parapseudid genera Discapseudes Băcescu and Guţu, 1975 and Halmyrapseudes Băcescu and Guţu, 1974 construct well–developed tubes (Băcescu and Guţu, 1974, 1975; R. Heard, pers. obser.). However, there can be different interpretations of whether or not members of the subfamily Kalliapseudinae are tube or burrow dwellers, or both. Based on the authors’ personal observations and those of Drumm (2005), we consider M. macsweenyi to be a burrow–lining that remains intact. Members of the Kalliapseudinae appear to be suspension or filter feeders occupying permanent domiciles in soft–bottom substrata (e.g., sand, sand–silt, mud). In contrast, members of the subfamilies Hemikalliapseudinae and Tanapseudinae , which are also known from soft–bottom habitats, appear to be fossorial deposit feeders that lack permanent domiciles.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Tanapseudes sinensis Bamber, 2000
| DRUMM, DAVID T. & HEARD, RICHARD W. 2011 |
Tanapseudes sinensis
| Gutu, M. & Angsupanich, S. 2005: 51 |
| Bamber, R. N. 2000: 45 |