
Tlalocohyla celeste, Varela-Soto & Abarca & Brenes-Mora & Aspinall & Leenders & Shepack, 2022
|
publication ID |
https://doi.org/10.11646/zootaxa.5178.6.1 |
|
publication LSID |
lsid:zoobank.org:pub:DB0FBF00-47CE-4DD8-B239-BEA81464D38F |
|
DOI |
https://doi.org/10.5281/zenodo.7043938 |
|
persistent identifier |
https://treatment.plazi.org/id/E5316619-D55E-4C46-FF28-FCEBA642FE08 |
|
treatment provided by |
Plazi |
|
scientific name |
Tlalocohyla celeste |
| status |
sp. nov. |
Tlalocohyla celeste sp. nov.
Tapir Valley Tree Frog, Rana Arbórea del Valle del Tapir
( Figures 2–7 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 )
Holotype. UCR 23700, an adult male from Costa Rica, Alajuela, Upala, Bijagua, Tapir Valley Nature Reserve (10.72˚ N, 85.01˚ W; 660 m asl), collected on September 13, 2021, by Juan G. Abarca, Valeria Aspinall, Donald Varela, and Esteban Brenes.
Paratopotypes. Two adult males, ECB-Anf-f50-08-01,01 and YPM 13222, same data as holotype; UCR 23701, an adult female collected by Donald Varela on September 15, 2021 .
Generic placement. Bayesian ( Fig. 1 View FIGURE 1 , Suppl. fig. 2) and Maximum likelihood analyses (Suppl. fig. 3) confirm the inclusion of the new species in the genus Tlalocohyla .
Diagnosis. The new species can be diagnosed by the following combination of traits: (1) small size (adults males: 21 mm, females: 24 mm of SVL; n=4); (2) tympanic membrane not-evident; (3) vocal sac developed, single, subgular, extending to the pectoral region; (4) vomerine teeth absent; (5) dorsal color yellow-green (#103); (6) ventral skin transparent; (7) white peritoneum covering all internal organs; (8) presence of an incomplete sulfur white (#96) dorsolateral stripe that originates at the posterior edge of the orbit and extends posteriorly to a point more than halfway towards the insertion of each hindlimb, bordered above for its entire length by a diffuse mahogany red (34) stripe that extends anteriorly towards the tip of the snout; (9) presence of a small light cyan (#158) axillary membrane.
Within its genus, Tlalocohyla celeste can be differentiated from T. godmani and T. loquax by the following characteristics (condition for T. celeste in parentheses): maximum SVL 45 mm ( 24 mm); venter pale lemon yellow or creamy yellow (transparent ventral skin) and extensive axillary membrane (very small axillary membrane). Tlalocohyla celeste is similar in overall appearance to T. smithii and T. picta , but can be differentiated based on the following characteristics: from T. smithii by its smaller size, maximum SVL 30 mm ( 24 mm); pale yellow, tan or brilliant yellow dorsum (bright yellow-green with mahogany red spots); a thin complete white or cream dorsolateral stripe that is usually bordered below by a thin brown line (an incomplete sulphur white dorsolateral stripe bordered above by a diffuse mahogany red stripe); white belly (transparent); vomerine teeth present (absent); and from T. picta by the relatively uniform yellowish tan dorsal coloration (bright yellow-green with mahogany red spots); presence of a complete white or cream dorsolateral stripe that may be bordered below by a poorly defined reddish, brown or gray band (an incomplete sulphur white dorsolateral stripe bordered above by a diffuse mahogany red stripe); white bones (green bones) and vocal sac yellow (brilliant light cyan). Tlalocohyla celeste also differs from these two species in its advertisement call and tadpole morphology (see below).
Tlalocohyla celeste is among the smallest hylids in Costa Rica, slightly smaller than Isthmohyla zeteki , and may be confused with other small hylid species present within its range. Members of the genus Dendropsophus are small tree frogs with an axillary membrane but none are green in coloration with transparent ventral skin. Scinax species have a similar, rounded protruding lateral snout profile but are significantly larger in size and never have light dorsolateral markings. Tlalocohyla celeste is most likely to be confused with juveniles of the syntopic Boana rufitela , but the latter has a rounded snout in lateral profile and its white dorsolateral lines are complete, extending from the snout tip backwards to the vent, passing over top of the hindlimbs. These light markings fade and eventually disappear completely as young B. rufitela mature. Because of its bright green coloration and transparent ventral skin, T. celeste may be mistaken for a glass frog (family Centrolenidae ), however glass frogs invariably lack a pattern of light dorsolateral stripes.
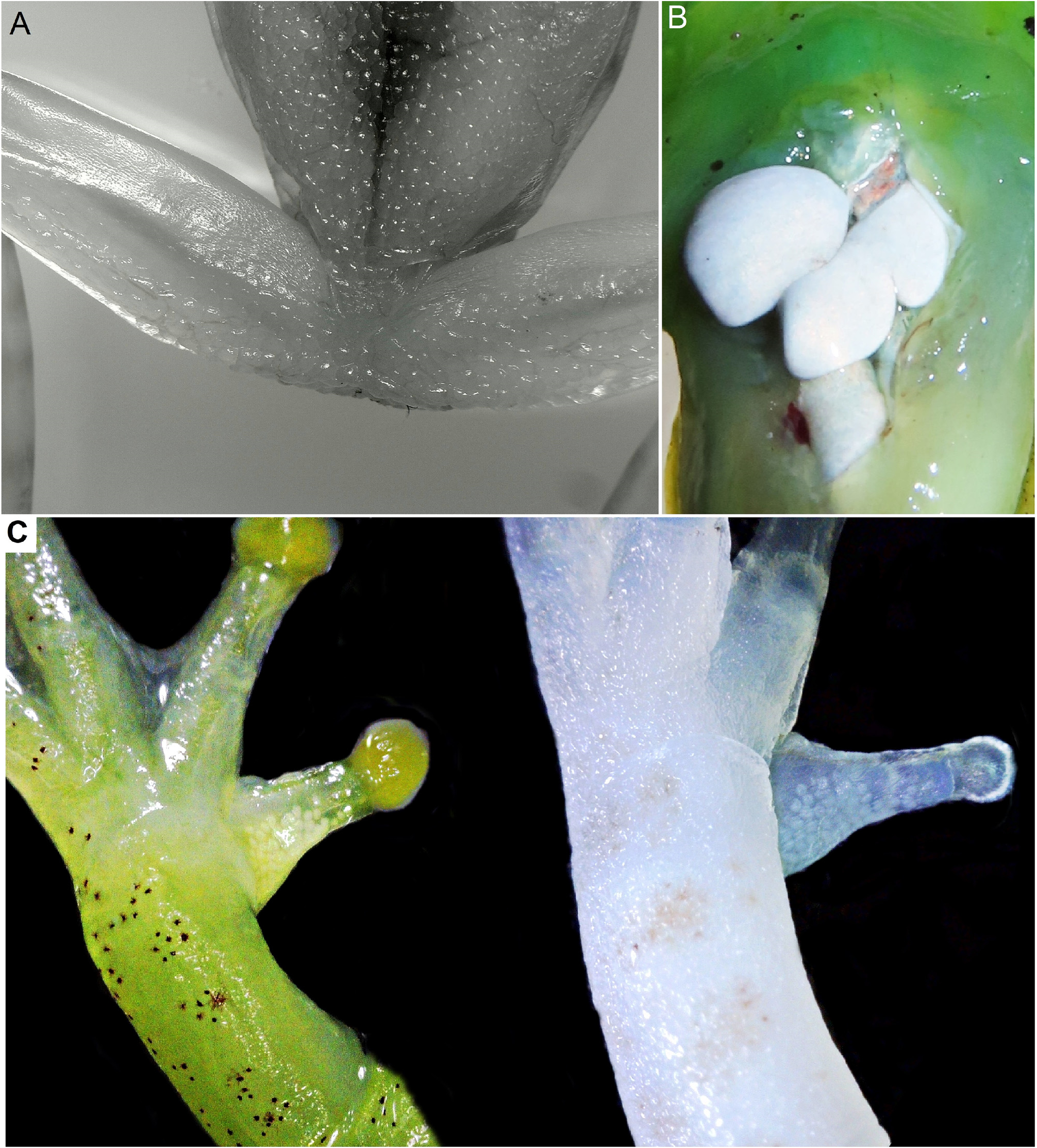
Description of holotype. An adult male with a SVL of 20.10 mm ( Fig. 2 View FIGURE 2 ), in a good state of preservation, head slightly longer than wide, HL 38.3%, and HW 36.5% of SVL. Snout nearly rounded in dorsal view, rounded protruding in profile. Nostrils rounded, dorsolateral, directed laterally, located on slightly raised processes, IND 25.8% of HW. Canthus rostralis well defined and rounded. Loreal region concave. Eyes large and protuberant, ED 39.1% and 10.7% of HW and SVL, respectively. Pupil horizontally elliptical. Tympanum small, TD 23.0% of ED, tympanic membrane inevident, but tympanic annulus visible through the skin ( Fig. 3C View FIGURE 3 ), tympanum matches character combination number 2 in Lynch and Duellman (1997). Tongue ovoid, free laterally and posteriorly. Texture of tongue granular. Premaxillary and maxillary teeth present, no vomerine teeth visible between choanae. Choanae rounded anteriorly and widely separated. Vocal apertures are slit-like and elongated, nearly parallel the mandible. Vocal sac single and subgular, extends posteriorly to the pectoral region ( Fig. 2B View FIGURE 2 and Fig. 7 View FIGURE 7 ).
Forelimbs not hypertrophied, upper arms and forearms slender, consistent throughout in its thickness. Hands proportionally large, HAL 33.1% of SVL. Lacks ulnar skin fold. A small axillary membrane present. Fingers robust, relative finger length I<II<IV<III. Discs moderately expanded and round (Fin3DW 25% of ED). Finger discs are comparable in size but finger disc I is slightly smaller than others. Subarticular tubercles single, rounded or globose, large and slightly bifid in fingers III and IV. Supernumerary tubercles large, single and rounded. Accessory palmar tubercles numerous, rounded, large and globular; span the entire palmar area in a tightly packed way. Inner metacarpal tubercle (thenar) elongated, elliptical, globular and large; outer metacarpal tubercle (palmar) not clearly differentiated from the accessory palmar tubercles. White smooth nuptial pads without epidermal projections, at the base of the first finger, formed by a grouping of glandular acini, each circular or ovoid in shape ( Fig. 6C View FIGURE 6 ). The interdigital membrane between fingers I–II is vestigial, but it is slightly more developed between fingers II–III–IV. Webbing formula I3+–2- II 2-– 3-III3- –2-IV.
Hind limbs slender, THL slightly greater than FL and slightly smaller than TL. TL of about 57.0% of SVL, FL 43.5% of SVL. Toes robust. Relative length of toes I<II<III<V<IV. Discs rounded and expanded. Subarticular tubercles single, rounded and globular. Supernumerary tubercles small, single and rounded. Inner metatarsal tubercle medium-sized, elliptical; outer metatarsal tubercle absent. Webbing formula I 2 - –2 + II 11/2 –3 + III 11/2 –3 - IV 2 + – 11/2 V, free parts of toes fringed. Cloacal dermal fold absent.
Body covered in smooth skin throughout, except for the central region of the venter and the undersides of the thighs where skin texture is distinctly granular ( Fig. 6A View FIGURE 6 ).
Measurements of the holotype (in mm). SVL 20.10; HL 7.70; HW 7.35; IND 1.89; IOD 3.26; ED 2.14; EN 1.88; TD 0.49; HAL 6.65; FL 8.75; TL 11.45; Fin3DW 0.55; Toe4DW 0.75.
Coloration of the holotype in life (nocturnal coloration). Background color of head, upper back and limbs brilliant yellow-green (#103), with numerous mahogany red (#34) blotches relatively evenly dispersed over the dorsal surfaces of the head, body and lower limbs ( Fig. 4A–B View FIGURE 4 ). A narrow sulfur white (#96) dorsolateral stripe starts behind each eye and extends towards the insertion of each hind limb, gradually fading out near a point about halfway down the body. A relatively poorly defined mahogany red stripe extends from the tip of the snout along the canthal region, and continues post-ocularly following the upper border of the sulfur white dorsolateral line; it gradually fades out at the same point as the light stripe. Lateral surfaces of the body and head, below the dorsolateral lines, lack mahogany red spots. Upper surfaces of finger and toe tips light greenish yellow (#87). Upper edge of orbit Pratt’s rufous (#72); remainder of upper eyelid suffused with spectrum yellow (#79). Posterior and anterior edge of orbit bordered by a narrow white line. Iris gold with raw umber (#23) reticulations. Vocal sac yellow-green (#103) when deflated, turquoise green (#147) when inflated. Skin covering ventral surfaces transparent, allowing muscles, veins, bones, and intestines to be visible ( Fig. 5C–D View FIGURE 5 ). Internal organs black, but hidden from view by an opaque, white peritoneum ( Fig. 6B View FIGURE 6 ).
Coloration of the holotype in preservative. In formalin, yellow-green dorsal coloration becomes pale greenish white (#97), while mahogany red markings (dorsolateral lines and dorsal spots) turn pale vinaceous (#247). The color pattern fades in a similar fashion after ethanol preservation, leaving dorsolateral markings and dorsal spots faded but visible ( Fig. 2 View FIGURE 2 ). The iris becomes black ( Fig. 3 View FIGURE 3 ). Hands, feet, and tips of digits fade to a uniform creamywhite. Skin on ventral surfaces retains some transparency in preservative.
Variation. Morphometric variation is detailed in Table 1 View TABLE 1 . Females are larger than males ( Fig. 4C View FIGURE 4 ) and lack nuptial pads, vocal slits and a vocal sac. The paratopotypes are comparable in coloration to the holotype. There is a noticeable difference between nocturnal ( Fig. 4D View FIGURE 4 ) and diurnal coloration ( Fig. 4F View FIGURE 4 ) in this species, especially in the intensity and visibility of the mahogany red dorsal spots and dorsolateral lines. These markings are less prominent during the day, causing the dorsal pattern to appear more uniformly green and less spotted. In addition, the eye color brightens during the daytime to a lighter silver-gray ( Fig. 4F View FIGURE 4 ). The presence of blue green bones, lymph, and other tissues ( Fig. 5 View FIGURE 5 ) indicates that this species has physiological chlorosis, a trait that has been documented in other arboreal frogs ( Taboada et al. 2020).
Metamorphs and small juveniles exhibit a lighter green dorsum and lack the light dorsolateral stripes during early developmental stages ( Fig. 11B–C View FIGURE 11 ). The small, light cyan axillary membrane ( Fig. 5F View FIGURE 5 ) is not seen in juveniles. Such young individuals generally display clusters of mahogany red blotches on the dorsum, most densely concentrated anteriorly but with considerable variation in their aggregation patterns. A broad, dark canthal stripe and short postocular stripe are usually present in juveniles, while the dorsal surfaces of the head, body, and limbs are marked with bold dark brown to purplish-brown spots. Dark-colored spots and stripes lighten ontogenetically likely to transform into the adult color pattern as the frogs age ( Fig. 4G View FIGURE 4 , 5E View FIGURE 5 and 11D View FIGURE 11 ).
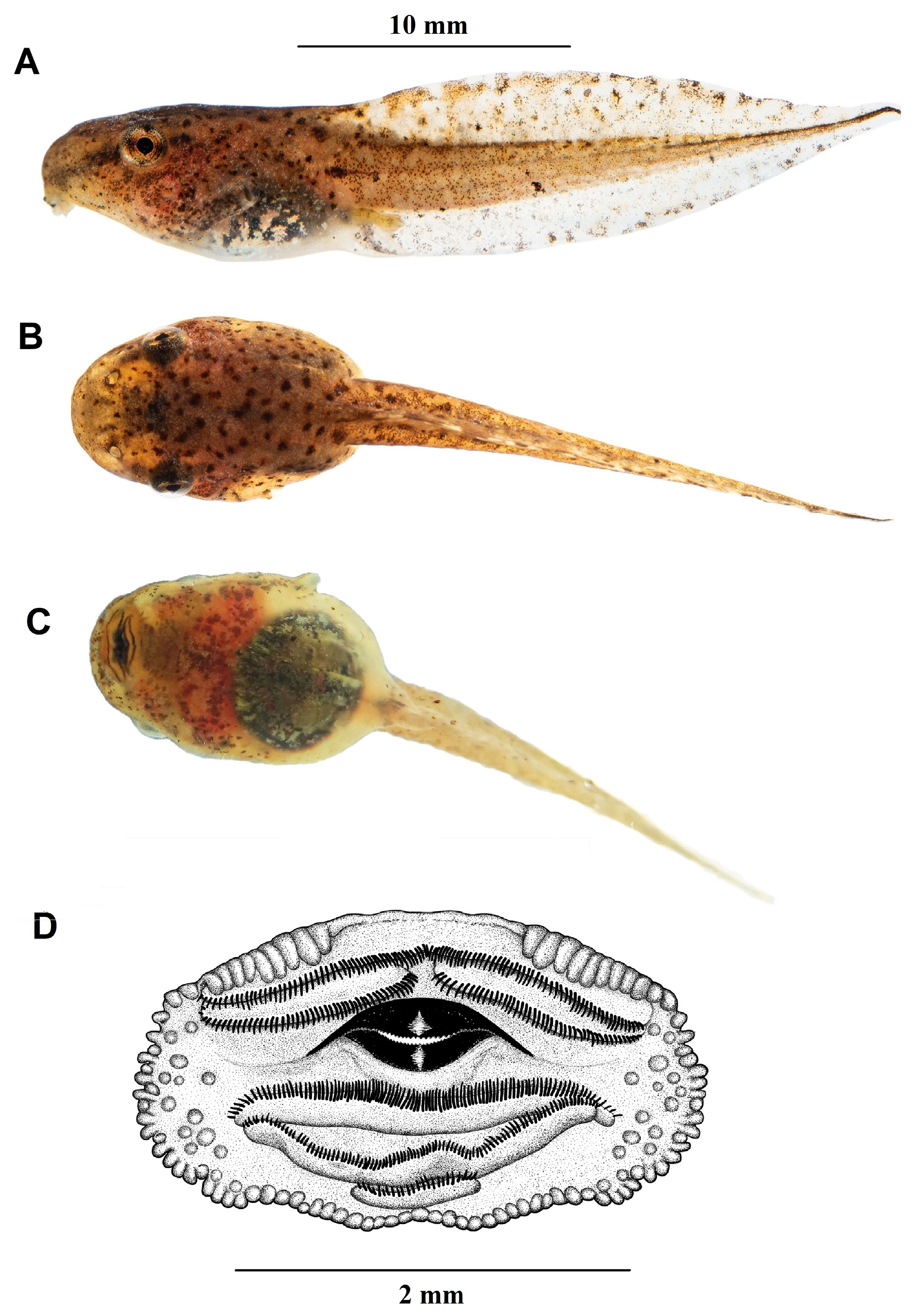
Tadpole description. Tadpoles of Tlalocohyla celeste ( Fig. 11A View FIGURE 11 and Fig. 12 View FIGURE 12 ) in Gosner stages 33–44 conform to the following description: body depressed (BH/BW=0.87–0.94), ovoid in dorsal view and ovoid or elliptical in lateral view, from 0.35-0.40 times the TL. Snout rounded in dorsal and lateral aspects. Eyes medium-sized (ED/ BL=0.11–0.16), laterally located, visible in ventral view. The nostrils are situated approximately midway between the eyes and the tip of the snout. Nares rounded, medium-sized (ND/BL=0.03–0.05), dorsally located and dorsally directed. Well-developed fleshy flanges on the marginal rim. Spiracle sinistral, lateral, directed posterodorsally, and located on the posterior third of the body. Spiracle opening small (SL/BL=0.13–0.16). The intestinal mass is positioned perpendicular to the longitudinal body axis and visible through the clear ventral skin. The cloacal tube is short, dextral and has a triangular apex.
The small oral disc (ODW/BW=0.39–0.46) is positioned anteroventrally. Oral disc not emarginated; has a single row of short marginal papillae with a small anterior gap. Several submarginal papillae are randomly distributed laterally in the oral disc ( Fig. 12D View FIGURE 12 ). LTRF 2(2)/3; A-1 and A-2 of equal size; A-2 with a narrow medial gap; P-1 equal in size to P-2; P-3 approximately one quarter the size of P-1 and P-2. Jaw sheaths small, measuring less than half of the oral disc; finely serrated along their entire margin. Anterior jaw sheath smoothly arched, posterior jaw sheath shallowly V-shaped. Tail of intermediate height (MTH/TAL=0.21-0.38) with moderately robust musculature (TMH/ BH=0.50-0.53); dorsal fin higher than ventral fin (DFH/VFH=1.20-1.56). Dorsal and ventral fins of intermediate height (DFH/TAL=0.07-0.11; VFH/TAL=0.05-0.09); dorsal fin initiates on the body at a point slightly anterior to the base of the tail; maximum fin height is achieved near the midpoint of the tail’s length. Ventral fin originates towards the end of the venter, anterior to the cloacal tube. The lateral line system is indistinct.
In life, the general color pattern of the body and tail musculature is pale cream, pale white or pale yellow, with small brown flecks on the dorsal surfaces ( Fig. 11A View FIGURE 11 – 12 View FIGURE 12 ). Dorsal surfaces of head yellow ochre (#14) to tawny olive (#17); lateral surfaces slightly lighter in hue, marked with concentrations of mahogany red (#34) to warm sepia (#40) spots. A pattern of relatively evenly-spaced, rounded warm sepia (#40) spots is a prominent feature of the dorsal surfaces; dorsal and lateral surfaces also display a fine dusting of irregular white or gold specks. A distinct, but poorly defined warm sepia (#40) stripe extends from a point near the anterior edge of the oral disc towards the eye and continues as an indistinct postocular stripe onto the body. Iris gold, marked on either side of the pupil with a warm sepia (#40) spot; both dark spots align with the dark eye stripe that originates at the snout tip. Ventral skin is transparent, revealing reddish hues anteriorly at the thoracic level and cream white (#52) coloration at the level of the intestines. Throughout the larval development period, the intestinal tract becomes increasingly suffused with black pigment. This ontogenetic change creates a striking black-and-white ventral pattern that contrasts sharply with the dorsal and lateral coloration. Tail fins are transparent, marked with black or raw umber (#23) blotches of irregular shape and size, as well as small white spots. Tail musculature is yellow ochre (#14) with black or warm sepia (#40) spots, occasionally with a fine black line on the medial side of the tail that can be intact or interrupted. Upon preservation in 10% formalin, the larval coloration fades to pale greenish white (#97), whereas the eye stripe, dorsal spots and tail markings become black.
Tadpoles of Tlalocohyla celeste differ most notably from other tadpoles in the genus as follows (Gosner stages in parenthesis): in tadpoles of T. godmani (38), T. loquax (26), and T. picta (38), the dorsal fin originates on the body at the level of, or anterior to the location of the spiracle ( Lee 1996; Duellman 2001), whereas in T. celeste the dorsal fin originates at the base of the tail. Tadpoles of T. smithii (37) are most similar to those of T. celeste as their dorsal tail fin starts close to the tail base. However, tadpoles of T. smithii differ in having the posterior third of the tail black and the height of their dorsal fin is relatively constant throughout until it tapers to a fine tip ( Caldwell 1986). Tadpoles of T. celeste lack a black tail tip and their dorsal fin increases in height towards the middle of the tail before tapering towards the tip.
Advertisement call. The vocal repertoire of Tlalocohyla celeste is composed of at least three call types. Call type I ( Fig. 8A–C View FIGURE 8 ) is composed of a trill of notes, with average call duration of 0.34 s (0.30– 0.39 s; n=17), emitted at intervals of 2–10 s (n=14) and average call repetition rate of 12 calls/minute. Call type I is composed of 9–13 notes, each note having average duration of 0.01 s (0.01– 0.02 s; n=17), with a note repetition rate of 33.4 notes/s and inter-note interval of 0.01 s (0.01– 0.02 s; n=17). Notes that comprise the call’s trill-pattern can be modulated to increase amplitude towards the end of the call or display similar amplitude throughout ( Fig. 8A View FIGURE 8 ). Call dominant frequency ranges 3.80–4.30 kHz ( Fig. 8D View FIGURE 8 ). Bandwidth values range from 1.13 (0.90–1.40) to 13.0 (10.3–23.0) kHz (n=17). This call has several sparse harmonics and each pulsed note (30–50 pulses per note) displays an initial high amplitude pulse but subsequently decreases to lower amplitude pulses ( Fig. 8E View FIGURE 8 ).
Call type II ( Fig. 8F–I View FIGURE 8 ) is composed of one to three squeak-like modulated pulsatile notes, each note having an average duration of 0.08 s (0.07– 0.10 s; n=4), with a note repetition rate of 3.4 notes/s and inter-note interval of 0.02 s (0.01– 0.02 s; n=4). The dominant frequency ranges from 4.00–4.40 kHz, bandwidth values range from 0.85 (0.40–1.5) to 12.1 (11.2–14.6) kHz (n =4). Call type II usually was emitted 0.17– 0.21 s (n = 4) after call type I.
Call type III ( Fig. 9A–D View FIGURE 9 ) is a croak of pulsed notes with an average duration of 0.29 s (0.21– 0.34 s; n=7), emitted at intervals of 0.35– 1.61 s (n=7), with an average call repetition rate of 23 calls/s (n=7). Call type III is composed of 3–5 pulsed notes of modulated amplitude, with an average call duration of 0.29 s (0.21– 0.34 s; n=7). Each note has an average duration of 0.03 s (0.03– 0.04 s; n=7), an inter-note interval of 0.04 s (0.03- 0.05 s; n=7), and 60–90 pulses per note. The dominant frequency ranges from 3.70–4.10 kHz; bandwidth values range from 1.39 (1.10–1.6) to 11.7 (11.1–13.3) kHz (n=7). Call type III can be emitted alone, after, or preceding call type I.
During periods of breeding chorus activity, males have been observed emitting a) only call type I, b) one to three type II calls, followed by a single type I call, c) emit several calls of type III followed or preceded by call type I, or d) produce all three call types sequentially ( Fig. 9E View FIGURE 9 ). Call type I of T. celeste is more similar to the trills produced by T. picta and T. smithii but can be differentiated by the following characteristics (condition for T. celeste in parentheses): Call type I differs primarily in the number of notes per call: the trill speed is faster in T. picta = 60–70 notes/s and T. smithii = 43–51 notes/s than in T. celeste (25–37 notes/s; n=17). In addition, T. picta and T. smithii have a higher dominant frequency, 4.6–6.30 kHz and 4.29–6.64 kHz respectively, when compared with T. celeste (3.80–4.30 kHz, n = 28).
Reproduction. Males of Tlalocohyla celeste have been observed calling year-round but display a significant increase in calling activity during periods of heightened precipitation. Vocalizations are generally first observed around 16:00 h and continue throughout the night, until sunrise. Individuals can be heard during the day, especially during periods of heavy rainfall. Most calling activity takes place along the vegetated wetland perimeter, density of calling males decreases towards the center of the wetland where water levels are deeper. Males have been observed calling in a head-down, horizontal, or head-up position, as they perch on vegetation. Individuals occupied a variety of herbaceous plant perches, but most were found on the locally common grass Rhynchospora corymbosa (Cyperaceae) ( Fig. 7 View FIGURE 7 ). Calling males are positioned 0.1–3 m above the ground, either calling in isolation or in small groups, with individuals often perched as little as 15–20 cm from one another. Satellite male behavior has been observed in this species, a trait also reported for T. picta ( Roble 1985) .
Females of this species, detected less frequently than males, were observed active in the periphery of the wetland and moved around calling males. On several occasions, females were found in the interior of the forest that surrounds the wetland, some 20 m from the water’s edge. Gravid females were first observed on June 12th, 2021 ( Fig. 4E View FIGURE 4 ), followed by subsequent sightings through September.
Tlalocohyla celeste exhibits axillary amplexus ( Duellman & Trueb 1986) ( Fig. 4C View FIGURE 4 and 10A View FIGURE 10 ). The first amplectant pair was discovered on June 13th, 2021 at 5:00 h. Amplexus and oviposition have been observed at night and during the pre-dawn morning hours. The amplecting pair separates immediately after oviposition.
All egg masses observed to date have been deposited at the tip, or along the drooping edge of leaves located directly above the water ( Fig. 10 View FIGURE 10 ). This indicates that T. celeste employs reproductive mode 24 ( fide Haddad & Prado 2005) where arboreal eggs hatch into exotrophic tadpoles that drop into lentic water. Egg masses contain 20–61 unpigmented eggs (n=8) ( Fig. 10B–D View FIGURE 10 ). Measurements taken from three different egg masses indicate that each clutch occupies an area between 12–22 mm in width and 23–28 mm in length; eggs measure an average of 2.0 mm in diameter (without the clear jelly capsules). One gravid female maintained in captivity for two days deposited three clutches containing 23, 45, and 61 eggs respectively, indicating that females can engage in multiple oviposition events during each breeding cycle.
Early embryos, observed five days after oviposition, were uniformly white with visible external gills ( Fig. 10E View FIGURE 10 ). Seven days after oviposition, these embryos were light brown and marked with small black spots, particularly on the tail; the ventral region remained white. Nine days after oviposition, the embryos were brown with additional spots present on body and tail ( Fig. 10F View FIGURE 10 ). The tadpoles hatched ten days after oviposition.
Egg masses were observed from June to November. Metamorphs and small juveniles were observed between November and June. Based on observations derived from tadpoles reared in ex-situ conditions, larval development appears to be slow. Three months after hatching, tadpoles reared under controlled conditions remained at Gosner stages 36–39 ( Gosner 1960). A recent metamorph was measured to have a SVL of 7 mm ( Fig. 11C View FIGURE 11 ).
The following anurans were observed to occur syntopically and likely share resources with T. celeste : Agalychnis callidryas , Boana rufitela , Dendropsophus ebraccatus , Dendropsophus microcephalus , Dendropsophus phlebodes, Incilius valliceps, Leptodactylus melanonotus, Lithobates forreri, Lithobates vaillanti, Rhinella horribilis, Scinax boulengeri , Scinax elaeochroa , Tlalocohyla loquax . All were confirmed to reproduce in the same wetland.
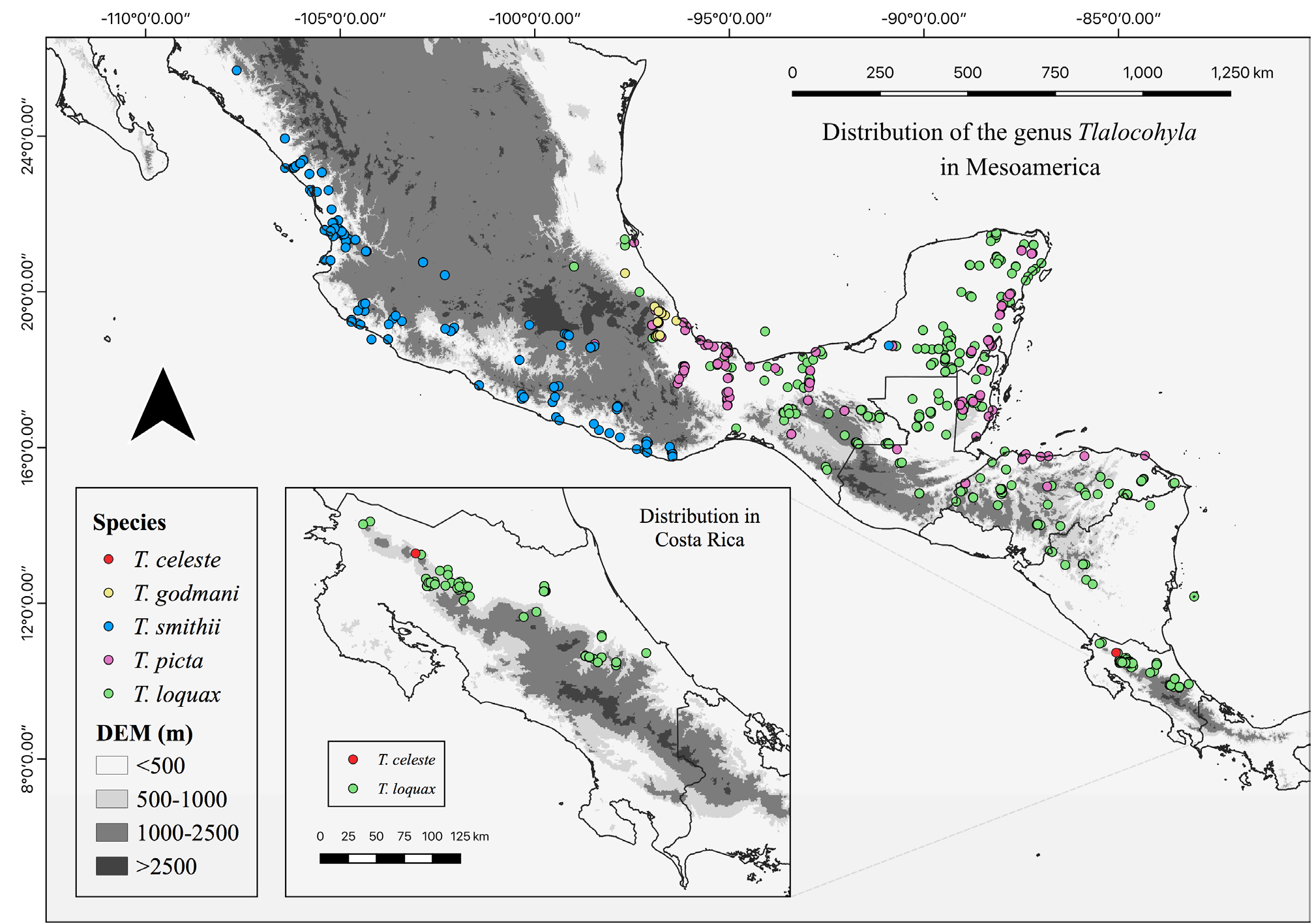
Geographic distribution and habitat. Tlalocohyla celeste is only known to inhabit a single wetland system in the privately owned Tapir Valley Nature Reserve (Lat: 10.71 N Long: - 85.01 W; elevation 660 m asl). The type locality is located between Tenorio and Miravalles volcanoes, in Bijagua de Upala, Alajuela Province, Costa Rica GoogleMaps ( Fig. 13 View FIGURE 13 ). The reserve’s life zones are classified as Tropical Premontane Wet Forest and Tropical Moist Forest ( Holdridge 1967). The number of dry months per year ranges from one to two, without a marked dry season. The mean annual precipitation is 3,500 mm and monthly temperatures ranges 20–33 ºC ( IMN 2021).
The frogs inhabit an 8-hectare marsh with a seasonally fluctuating, but permanent water level. The wetland is continually fed by a small tributary, by rainwater collected from surrounding hillsides, and likely by groundwater that wells up from bottom springs.A narrow discharge channel facilitates slow, directional water movement through the wetland. In the past, sections of this wetland have been channelized for irrigation purposes but it is now reverting to a more natural state. The surrounding hills and foothills of nearby mountains are largely forested, but areas of cultivated grassland are interspersed throughout the region. The shallow part of the marsh’s littoral zone is dominated by the grass Rhynchospora corymbosa (Cyperaceae) , whereas the riparian zone is occupied by a diverse assemblage of herbaceous vegetation ( Fig. 14 View FIGURE 14 ). Away from the water’s edge, the vegetation structure changes and herbaceous plants are replaced by woody shrubs and trees, which connect to forest in the upland areas.
Tlalocohyla celeste is an abundant species within the marsh at Tapir Valley Reserve. During periods of increased activity their calls can be heard throughout the littoral zone of the wetland, and one can readily observe up to 50 individuals in a single night. Long-term monitoring is ongoing to determine the population dynamics of this species. In spite of its local abundance, repeated visual and acoustic searches in nearby wetlands within a 2 km radius that seemingly displayed similar habitat quality and characteristics have thus far not resulted in the discovery of additional T. celeste populations.
Natural history. Individuals of this new species have been observed catching small flies and micro moths, and presumably its diet consists of a variety of small invertebrates. Although no direct observations of predation on adult T. celeste were made, its wetland habitat harbors abundant snakes that are known predators of anurans. In addition, ctenid spiders, which are common frog predators, abound. During the course of our surveys, opilionid arachnids were observed scavenging a dead frog and small wasps were seen attacking T. celeste egg masses and preying on early developing larvae
Etymology. The specific epithet “celeste” (light blue, or sky-blue in Spanish), is used as a noun in apposition, and refers to the striking blue coloration seen on this frog’s axillary membranes and groin area, as well as on the inflated vocal sac in adult males. Perhaps more importantly, the authors want to celebrate the river Río Celeste, famous for the distinctive turquoise-blue hue of its water, whose watershed feeds the wetlands that provide critical habitat for its namesake frog. Río Celeste represents a critical natural resource and an important economic driver for local communities, as well as a nexus for sharing the beauty of nature with visitors from outside the region. The fact that this frog was discovered through local wetland restoration efforts is a testament to the critical importance of protecting our aquatic resources.










| YPM |
Peabody Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.