
Tlaxcallicetus guaycurae, Cisneros, 2018
|
publication ID |
https://doi.org/ 10.26879/746 |
|
publication LSID |
lsid:zoobank.org:pub:138AE090-E19F-442E-8BDF-13B8401B755E |
|
DOI |
https://doi.org/10.5281/zenodo.11062533 |
|
persistent identifier |
https://treatment.plazi.org/id/3B27C833-31FE-4DF7-9929-78E860A1EEF8 |
|
taxon LSID |
lsid:zoobank.org:act:3B27C833-31FE-4DF7-9929-78E860A1EEF8 |
|
treatment provided by |
Felipe |
|
scientific name |
Tlaxcallicetus guaycurae |
| status |
sp. nov. |
Tlaxcallicetus guaycurae , new species
Figures 3-8 View FIGURE 3 View FIGURE 4 View FIGURE 5
zoobank.org/ 3B27C833-31FE-4DF7-9929-78E860A1EEF8
Diagnosis. The autapomorphic characters that distinguish Tlaxcallicetus guaycurae from other known Oligocene archaic chaeomysticetes include: i) presence of an external occipital sulcus (new term) on the posterior middle part of the supraoccipital; ii) well-developed and deeper dorsal condyloid fossae; iii) well-defined supracondylar septum (new term that refers to the prominent elevation of dorsal surface on the middle line of the foramen magnum); iv) a prominent exoccipital posteriorly directed farther from the posterior edge of the occipital condyles; v) inflated and rounded squamosal prominence; vi) posterior end of basioccipital crests transversally aligned to the level of the straight postglenoid process; vii) posteriorly exposed ventral surface of the paroccipital process of the exoccipital; viii) postglenoid process at same level that the ventral edge of the exoccipital, lateral view; ix) half-moon shaped fenestra rotunda; and x) a flattened anteroposteriorly compound posterior process.
Etymology. The specific name guaycurae (Latinized) recognizes the ‘Guaycura’ indigenous group that lived throughout the municipality of La Paz, Baja California Sur, region where the holotype was found.
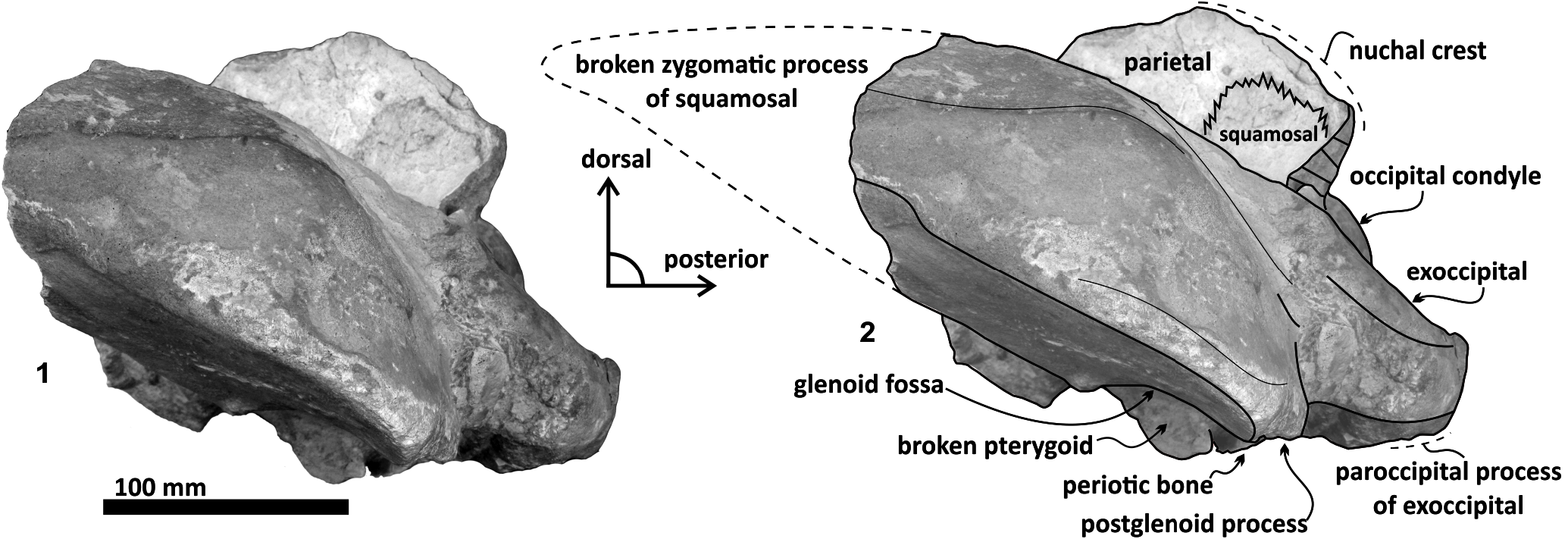
Material. Tlaxcallicetus guaycurae is known only from the holotype, MHN-UABCS_EcSj5/06/31 , which includes a partial cranium without rostrum, mandibles and postcranial bones. The best-preserved fragments of Tlaxcallicetus guaycurae include a left broken zygomatic process of squamosal, a well preserved left squamosal fossa, a glenoid fossa, a supraoccipital without apical portion, occipital condyles, left part of the exoccipital, basioccipital, and an almost complete left periotic bone. Specimen was collected by G. Gonzalez-Barba, December 1, 1991.
Type locality. El Saladito (mine spoils), San Juan de la Costa , La Paz, Baja California Sur, Mexico (24.43916 N, - 110.69512 W). See details in Figure 1 View FIGURE 1 GoogleMaps .
Horizon. From a phosphatic grey fine sandstone bed ~ 1 m above the Humboldt bed at the general stratigraphic column ( Figure 2 View FIGURE 2 ) of San Juan de la Costa ( Schwennicke, 1994).
Age. Late Oligocene, ~27 Ma based on the planktonic foraminifera (see above).
Description
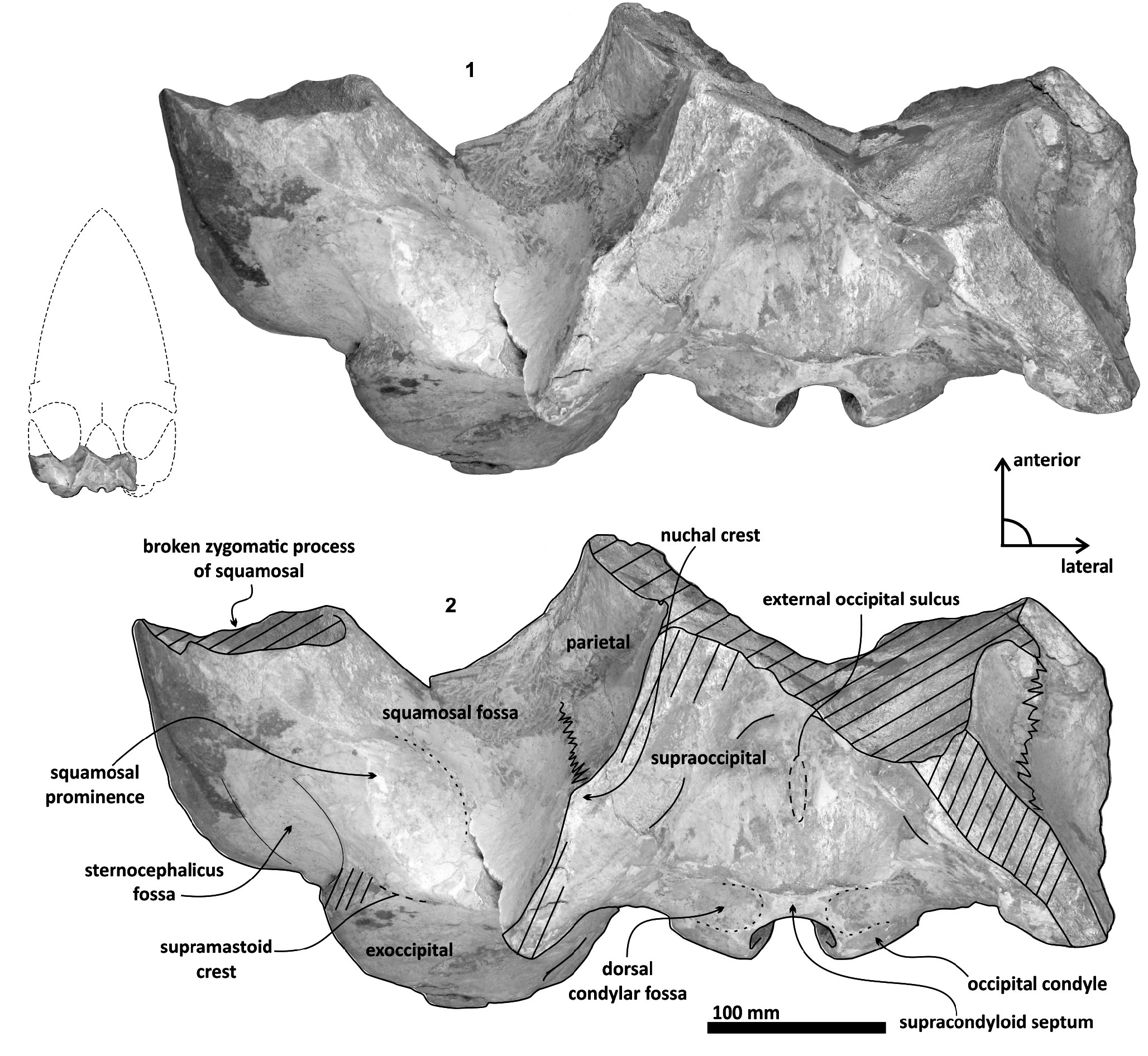
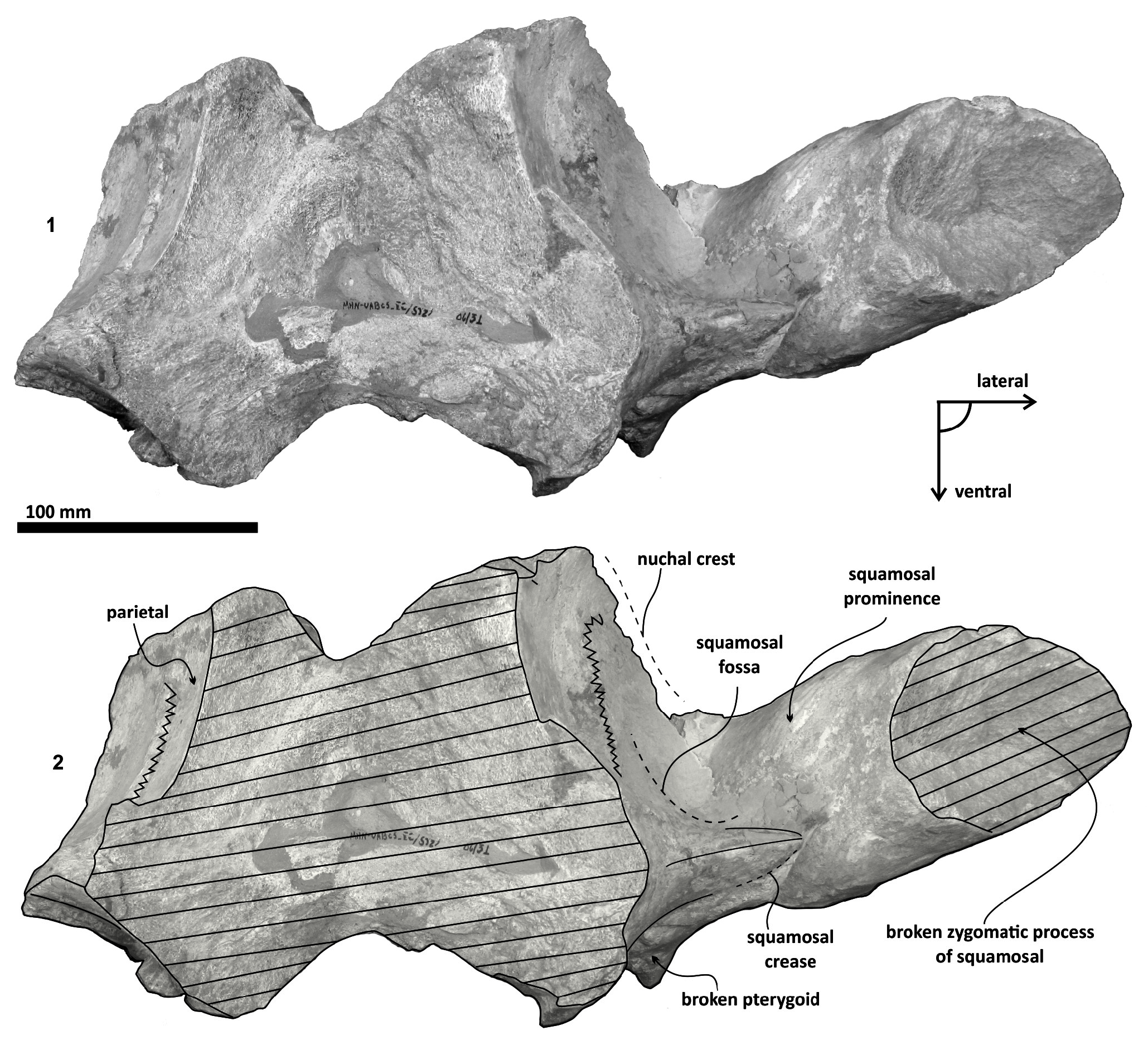
Parietal. A small posterior part of parietals is preserved. The surface is concave and fused to the squamosal. No foramina or other important characters are seen ( Figures 3 View FIGURE 3 , 5 View FIGURE 5 , 7 View FIGURE 7 ). The parietal/squamosal suture obliquely descends over the anterior portion of the squamosal fossa. Its contact with the sphenoid bone is not well defined; the suture is dorsally and posteriorly convex, close to the nuchal crest, and apparently, the parietal forms most of the nuchal crest together with the supraoccipital.
Supraoccipital. The apex of the supraoccipital is broken, and the nuchal crests are practically eroded. A small preserved anterior portion suggests that the anteromedial surface of the supraoccipital was elevated relative to the posteromedial part, and may have had a triangular profile in dorsal view ( Figure 3 View FIGURE 3 ). The nuchal crest is dorsally convex and ventrally concave on the parietosquamosal surface, and overhangs the squamosal fossa. Its posterior part turns down over the exoccipital/squamosal suture, and an inflexion defines the origin of the eroded supramastoid crest, which extended laterally over the exoccipital/squamosal contact. It does not follow a continuous path over the dorsal surface of the zygomatic process. The nuchal crest runs posteriorly directed and falls into a vertical profile as in the basilosaurids. The posterior middle part of the supraoccipital is depressed ( Figure 6). Its surface is smooth and flat (slightly convex) with a sagittal oval sulcus (external occipital sulcus) in the middle anterior part of the flat surface. The external occipital sulcus is around 5 mm deeper, and is unique for Tlaxcallicetus guaycurae ( Figure 9.3 View FIGURE 9 ). No other known mysticete has one. This sulcus is probably a reinforcement of the epaxial muscle for feeding proposes maintaining a strong atlantooccipital joint associated to the rectus capitis dorsalis major and the rectus capitis dorsalis minor (see Bouetel, 2005; Evans and de Lahunta, 2013).
Exoccipital. Posteriorly, the supraoccipital meets the exoccipital ( Figures 3 View FIGURE 3 , 6). Dorsal to the occipital condyles, a particularly deep, rounded, and well-developed dorsal condyloid fossa appears. This feature is present in other Oligocene primitive mysticetes as a shallow depression; for example, in eomysticetids ( Boessenecker and Fordyce, 2014), or some aetiocetids ( Barnes et al., 1995) but apparently, it is also present in many cetaceans and provides a room for movements of the atlas ( Mead and Fordyce, 2009). Between the condyloid fossae, a well-defined and prominent supracondylar septum is present. It is not certain if the latter is common in cetaceans with deeper dorsal condyloid fossae; however, a tenuous form of this septum is seen in some fossils (e.g., Eomysticetus whitmorei ). Laterally, the exoccipital is located posterior to the squamosal; it is prominent and posteriorly directed towards the posterior edge of the occipital condyles. This pronounced extension is dorsoventrally thick (~ 50 mm at its posterior end). From a dorsal view, it has a semicircular outline, and the dorsal surface is slightly concave. This feature is unique in Tlaxcallicetus . Ventrally, the paroccipital process in the exoccipital is thick (~ 30 mm anterposterioly) and is exposed posteroventrally. Apparently, its surface has a broad attachment area for the abductor digastric muscle (see El Adli and Deméré, 2015). The jugular notch is open widely between the exoccipital and basioccipital crest. A hypoglossal foramen is situated near to the posterior end of the basioccipital crest and opens into the jugular notch.
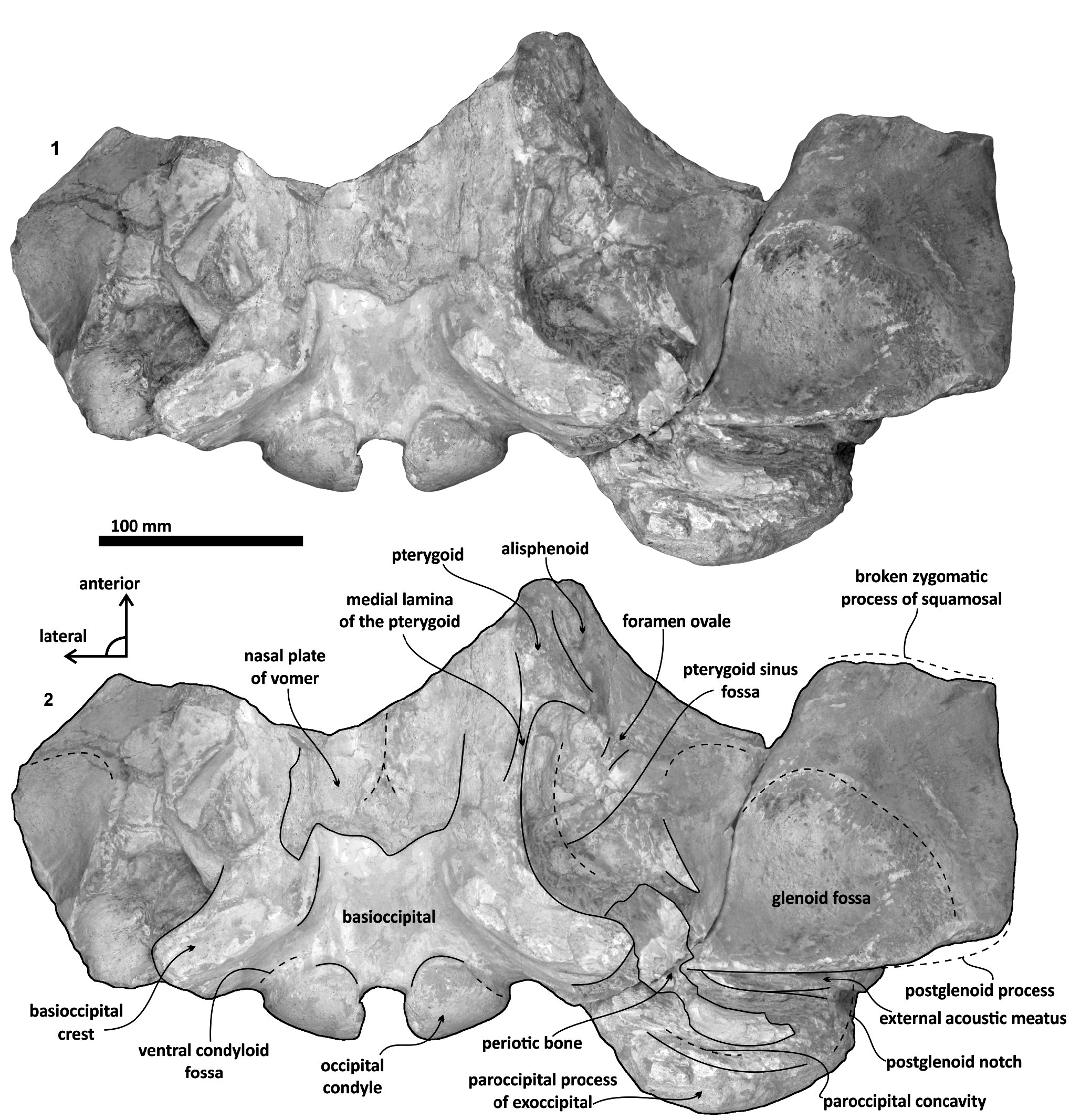
Basioccipital. The basioccipital forms the cranial base ( Figure 4 View FIGURE 4 )–basilar part (sensu Fitzgerald, 2010). It is smooth medially and becomes flat to slightly convex dorsally. Anteriorly, it is fused to the basisphenoid and ventrally overlaps to the posterior end of vomer (nasal plates fragment). Ventrolaterally, the basilar part extends forming a prominent and thick basioccipital crest with a slightly eroded convex surface. Basioccipital crest is long and bulbous, mediolaterally and dorsoventrally thickened, as in all mysticetes ( Fitzgerald, 2010). The crest curves laterally in a right angle at the level of the postglenoid process. The entire crest extends between the middle part of the pterygoid sinus fossa and postglenoid process. Its lateral margin is concave and forms part of the pterygoid sinus fossa, and the blunt apex marginally overhangs the head of the periotic as well as over the periotic fossa. Moreover, the crest is transversally thick, around ~ 35 mm, near its posterior tip end. This latter feature also distinguishes it from Tlaxcallicetus sp.
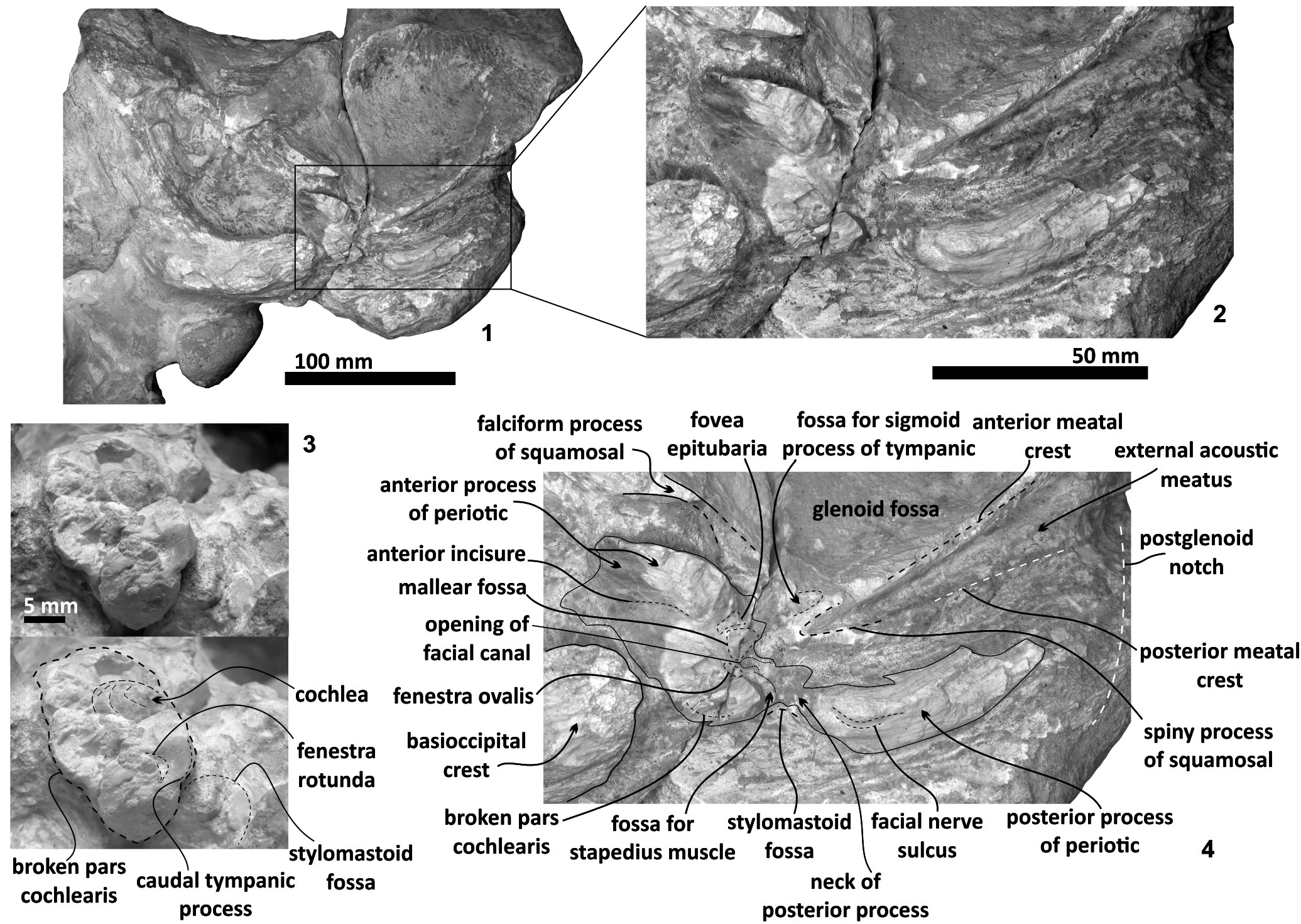
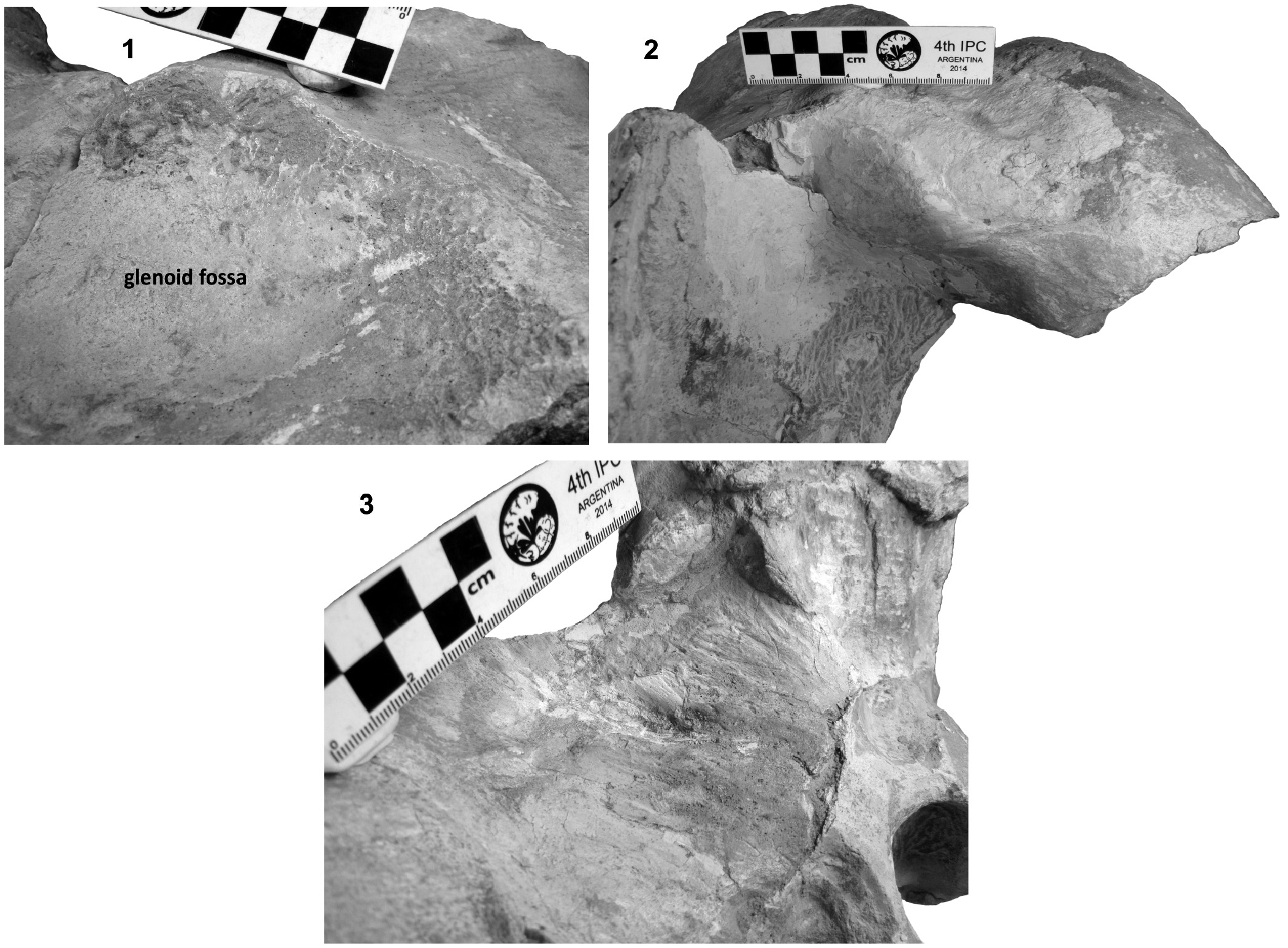
Squamosal bone. The dorsal squamosal surface meets the posterior part of the parietal, and both form the concave space of the squamosal fossa ( Figure 3 View FIGURE 3 , 5 View FIGURE 5 ). The fossa is anteroposteriorly short and transversally broad. A rounded and inflated squamosal prominence rules the lateral surface of the squamosal fossa at the medial part of the base of the zygomatic process of the squamosal. The base of the zygomatic process is transversally wide and posteriorly fused into the exoccipital. The supramastoid crest is broken, it is laterally directed and continues between the nuchal crest and exoccipital joint but is not present over the zygomatic process. Dorsolaterally and next to the squamosal prominence occurs a broad sternocephalicus fossa with a flat to slightly convex surface. Its oval area is limited by a blunt and thin ridge. Ventrally, the postglenoid process is exposed on the posterolateral margin of the base of zygomatic process. The apex of the zygomatic process is broken but in the anterior view can be seen a round transverse section that suggests a conical shape ( Figure 5 View FIGURE 5 ). The left glenoid fossa is broad and concave. Its outline is nearly semicircular with the anterolateral margin marked by a visible pitted surface ( Figure 9.1 View FIGURE 9 ). These pits could imply the presence of a possible non-synovial and fibrocartilaginous joint, similar to the one in balaenopterids ( Lambertsen et al., 1995). Posteriorly, the glenoid fossa is limited by a perpendicular and straight postglenoid process (relative to the sagittal axis), which is aligned to the rear of the basioccipital crest. The postglenoid process is relatively thick in lateral view and posteroventrally directed. Thus, it is different from the anteriorly curving postglenoid process present in archaic tooth-bearing mysticetes as the aetiocetids ( Barnes et al., 1995) or the eomysticetids ( Sanders and Barnes, 2002 a, 2002b, Boessenecker and Fordyce, 2015a, 2015b). Also, it is small and not prominent ventrally or posteriorly, and is aligned ventrally to the paroccipital process of the exoccipital. Posteriorly to the postglenoid process, a broad postglenoid notch opens. Inside, the external acoustic meatus is mediolaterally long and relatively narrow anteroposteriorly, between the anterior and posterior meatal crests of the squamosal. Medially to the glenoid fossa, the falciform process is broken. The pterygoid sinus fossa is a deep cavity with an ovoid shape and anterolaterally, the foramen ovale canal crosses the pterygoid sinus fossa. A small and indistinct fossa for sigmoid process is also present near the spiny process of the squamosal. The left periotic is in situ and practically complete within the periotic fossa.
Vomer/ Alisphenoid/ Pterygoid. Some fragments of the alisphenoid, vomer, and pterygoid are preserved. The pterygoid is lateral to the posterior end of the vomer (nasal plates) but is broken. Apparently, the pterygoid forms part of the foramen ovale, and a thin medial lamina of the pterygoid is preserved. Alisphenoid is fused laterally to the pterygoid and represents just a portion of the ventral surface of the braincase. The nasal plates of vomer are wide transversally and are divided by an eroded vomerine crest.
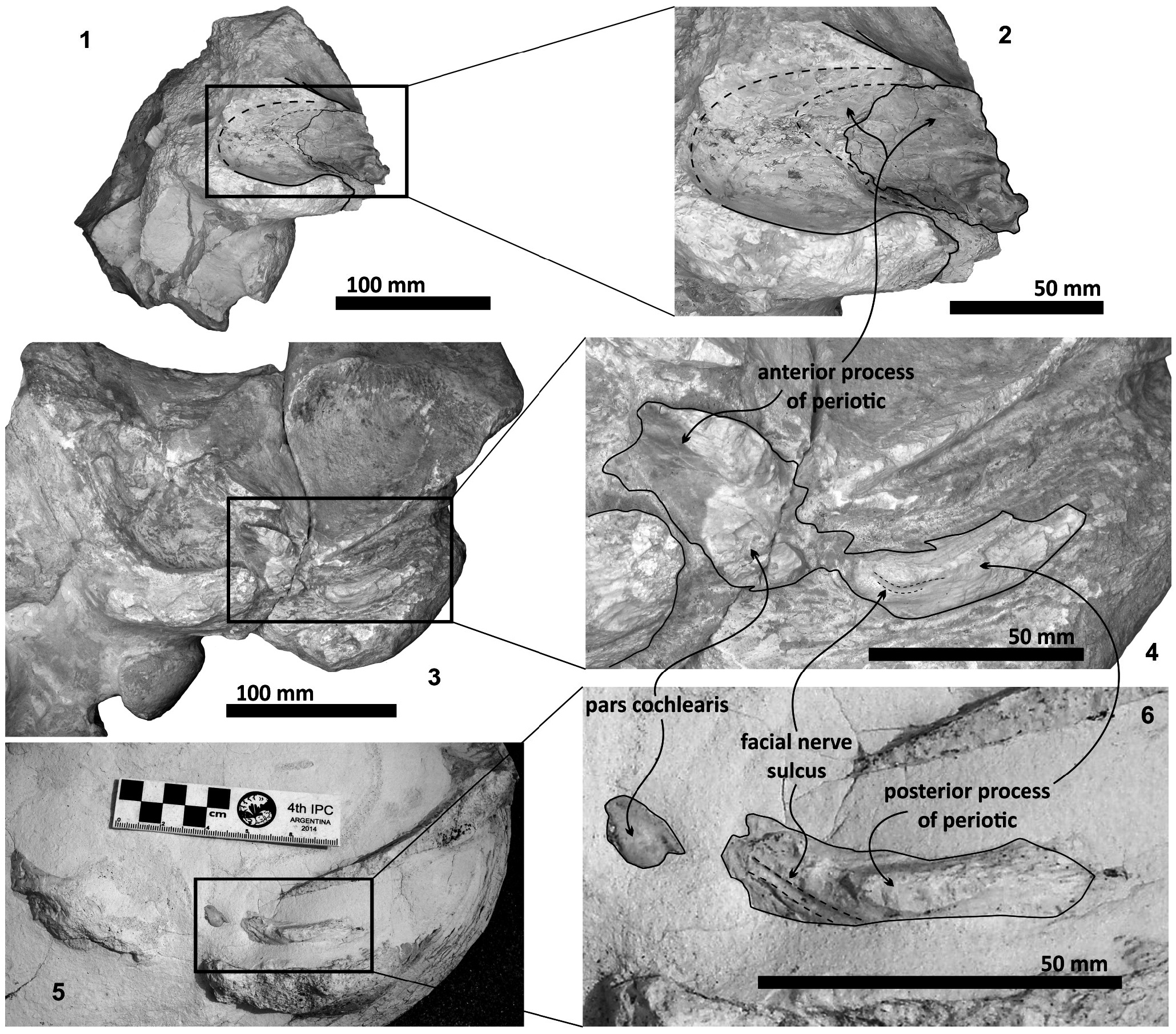
Periotic. The periotic is the best diagnostic feature of Tlaxcallicetus . In Tlaxcallicetus guaycurae , the periotic is basically complete, but the dorsal surface (internal acoustic meatus, suprameatal area, superior process, the aperture to the vestibular aqueduct, and the cochlear aqueduct) is not visible ( Figure 8 View FIGURE 8 ). However, it is anatomically similar to the Tlaxcallicetus sp. ( MU EcSj5/18/95), left periotic ( Figure 10 View FIGURE 10 ). The anterior process of the periotic is transversally narrow (~ 10.5 mm as maximum at the base near the fovea epituvaria), and very thin dorsally. The anterior keel is vertical. The anterodorsal angle and the anteroventral angle are broken. In the posteriomedial wall of the anterior process and anterior to pars cochlearis, a marked anterior incisure is present. Ventrally, the incisural flange is eroded. Posteriorly to the broken anteroventral angle, a rectangular and narrow fovea epitubaria appears. The adjacent mallear fossa is poorly preserved but looks rounded, concave with a well-defined margin, and oriented posteroventrally. The lateral tuberosity is not complete but the fragments suggest that it is blunt and indistinct; posteriorly, the fossa incudis is eroded. The fenestra ovalis is broken and nearly elliptical, oriented ventrally and located anteromedial to the distal opening of the facial canal. The fossa for the stapedius muscle is anteroposteriorly long, finishing in the stylomastoid fossa. The pars cochlearis is broken in its ventromedial surface, without evidence of an anterointernal angle or a median promontorial groove. The cochlea is exposed ventrally. Posteriorly, the fenestra rotunda is preserved as a half-moon groove and ventrally, the caudal tympanic process has a low profile with a blunt margin ( Figure 8.3 View FIGURE 8 ). The head of the periotic is connected by a cylindrical neck to the posterior process. On the dorsomedial surface between the posterior part of the pars cochlearis and the posterior process, a prominent depression of the stylomastoid fossa is present. The tympanic and periotic posterior processes are fused and form a compound posterior process oriented in a right angle to the axis of the anterior process. It is flattened anteroposteriorly. Near the neck and closer to the posterior margin of the posterior process, a transversally short facial nerve sulcus appears.

| MU |
Midwestern University |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Mysticeti |
|
InfraOrder |
Chaeomysticeti |
|
Genus |