
Varichaetadrilus harmani ( Loden, 1979 ) Loden, 1979
|
publication ID |
https://doi.org/ 10.5281/zenodo.173402 |
|
DOI |
https://doi.org/10.5281/zenodo.6263523 |
|
persistent identifier |
https://treatment.plazi.org/id/03B8F716-FFF4-FFB1-7C7D-FEB3FD27741D |
|
treatment provided by |
Plazi |
|
scientific name |
Varichaetadrilus harmani ( Loden, 1979 ) |
| status |
comb. nov. |
Varichaetadrilus harmani ( Loden, 1979) n. comb.
( Figs 1 – 3 View FIGURE 1 View FIGURE 2 View FIGURE 3 )
Tubifex harmani : Loden (1979), 601–605, Fig. 1 View FIGURE 1 .
Tasserkidrilus harmani: Brinkhurst (1986) , 150–151, 253; Fig. (unnumbered, p. 150).
Description of new material
The intact mature specimens were 26 and 29 mm long, with 93 and 123 segments, respectively. The immature specimens measured up to 23 mm in length (averaging about 10 mm), with segment number up to 117. Frequently, an unsegmented growth zone, or a posterior regenerate occurred. In several specimens a thinner, regenerating anterior portion was observed, consisting of 2–10 segments when countable ( Fig. 1 View FIGURE 1 B,C). A short, crippled individual had only 16 original segments, but with both ends in a state of regeneration. The preserved worms were slim, with a diameter of up to 0.3 mm in the anterior portion, 0.4–0.5 mm in the genital segments, and gradually tapering to about 0.15 mm in the tail.
Prostomium bluntly conical, separated with a furrow ( Fig. 1 View FIGURE 1 A,D). Intersegmental furrows shallow, without additional rings. Body wall smooth and transparent, but thicker and more opaque in I–III.
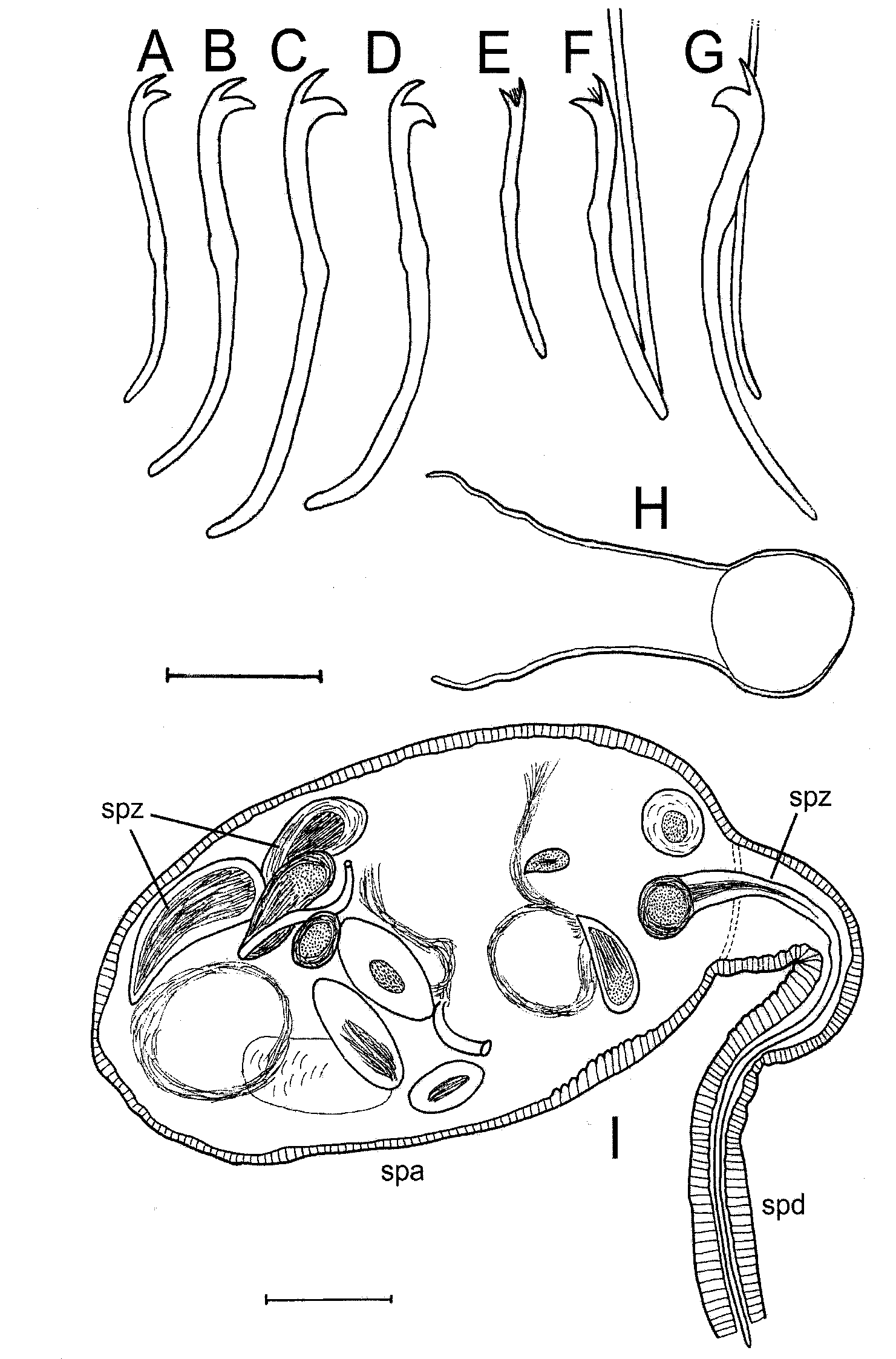
In anterior dorsal bundles, 1–2 very thin and smooth hair chaetae, mostly 300–400 μm long, and 2–3 slender pectinates, mostly 80–100 μm long, with equal teeth and several short intermediate denticles ( Fig. 2 View FIGURE 2 E,F). Anterior ventral bifids 3–4 per bundle, thicker (4–6 μm) and more curved than dorsal pectinates, mostly 80–100 μm long, with teeth of equal length but the upper one always thinner ( Fig. 2 View FIGURE 2 A–C). Both dorsal and ventral chaetae in II (and less so in III) considerably shorter than in the following segments ( Fig. 2 View FIGURE 2 A,E). Ventral chaetae of X and XI unmodified, in XI only two per bundle.
On midbody and tail, mostly two bifid crotchets per bundle, about 75–90 μm long, 6–7 μm thick and strongly curved, with the upper tooth much smaller than the lower one. Dorsal bundles usually contain also a thin hair chaeta, about 300 μm long ( Fig. 2 View FIGURE 2 D,G).
Clitellum in XI–XII ( Fig. 1 View FIGURE 1 A), 15–37 μm thick, mufflike except for a small median bare patch between and before ventral bundles in XI. Male pores close to ventral bundles in XI. Female pores on ventral chaetal line in 11/12. Spermathecal pores in X between ventral and dorsal bundles, slightly closer to dorsal ones. Ventral surface of genital segments in clitellate individuals deeply concave, fringed on both sides, with very broad and flat ventrolateral folds. Anterior ends of these folds ascend diagonally to dorsal chaetae in X; in XI–XII the folds proceed above ventral bundles, levelling gradually near 12/13. The folds are produced by contracting dorsoventral muscles and include some part of coelom with internal organs. All genital pores lie under the edge of the folds ( Fig. 1 View FIGURE 1 A, 3B,D).
Internal epithelium of body wall relatively thick in anterior segments. Ventral nerve ganglia with large lateral appendages in the posterior portion of V, VI and VII, and with smaller ones in VIII and XIV (not studied in posterior segments).
Pharynx thickwalled. Abundant pharyngeal glands reach the posterior side of 5/6. Chloragogen tissue on oesophagus beginning in VI. Intestine dilating either abruptly in X, or gradually; in mature individuals, considerable expansion only in XV ( Fig. 1 View FIGURE 1 A,D). Intestinal blood sinus weakly developed. No nephridia observed in preclitellar segments. No coelomocytes.
Dorsal blood vessel in anterior segments well developed, sometimes even wider than intestine (in IX–X of the sectioned specimen). Transversal vessels forming loops in IV–VIII. Those of IX transformed into short, thickwalled “hearts”, about 20 μm wide, and covered with their own chloragogen layer about 10 μm thick ( Fig. 3 View FIGURE 3 A,B).
Testes in X, small. Sperm sac reaching XV. Male funnels in X large and thinwalled, up to 180 μm high, flattened between spermatheca and body wall in the sectioned specimen ( Fig. 3 View FIGURE 3 B). Vasa deferentia shorter than atria (not exactly measured), directing almost straight backwards, joining smoothly the apical end of atrium. They are uniformly 30–35 μm wide, consisting of an internally ciliated, columnar epithelium, 4–10 μm high ( Fig. 3 View FIGURE 3 C,D).
Atria tubular, about twice longer than body diameter, with their proximal portion either winding in the dorsal half of XI, or reaching backwards into XII and with the distal portion inside ventrolateral body folds ( Fig. 1 View FIGURE 1 A, 3C,D). They are mostly 40–70 μm wide, but up to 80 μm near their proximal end; in cross sections, the distal end of one atrium tapered up to 25 μm, while the other remained wide until joining the penis. External muscle layer
2–8 μm thick, crossstriped with long transversal fibres throughout ( Fig. 1 View FIGURE 1 A). Internal epithelium 9 – 12 μm thick, often eroded, mostly forming longitudinal folds. The wider proximal end of atria can reveal thinner epithelium and more spacious lumen ( Fig. 3 View FIGURE 3 C,D).
No separate ejaculatory ducts. Solid prostate glands fill a large portion of coelom in XI–XII (but lacking in the anterior part of XI depicted on Fig. 3 View FIGURE 3 C,D), their short stalks joining the proximal, wider portion of atria subapically.
Penial sacs inside ventrolateral folds of XI, large, conical, slightly bent, with thin muscular wall, opening as simple male pores ( Fig. 3 View FIGURE 3 D). Penes much smaller than sacs, covered with distinct delicate (1 μm thick) cuticular sheath. The sheath is about 90 long, with a funnelshaped proximal end, 40–50 μm wide, tapering midpart, and abruptly expanding to a width of 30–45 μm at the distal end ( Fig. 1 View FIGURE 1 A, 2H).
Large ovaria in XI. Egg sacs can reach XIV ( Fig. 1 View FIGURE 1 A). Female funnels not seen.
Spermathecae in X, but in the sectioned specimen one ampulla was pressed backwards, into XII – XIII ( Fig. 3 View FIGURE 3 A–D). Ampulla roundish or oval, up to 400 μm long and 200–300 μm wide when filled, with the epithelial wall about 10 μm thick ( Fig. 2 View FIGURE 2 I, 3A). Distinctly separated duct of the same length or slightly longer than ampulla (up to 550 μm), 30–60 μm wide, covered with a layer of transversal musculature, 1–4 μm thick; its epithelium 12 – 14 μm thick and internally folded, with the lumen thus appearing starlike in cross sections ( Fig. 3 View FIGURE 3 C). Spermatozeugmata numerous, about 450 μm long; their spindleshaped main part 80–105 μm long and up to 47 μm wide, while a much longer tail narrows gradually from 20 to 7 μm ( Fig. 2 View FIGURE 2 I, 3A).
Comparison with the original description and type material
The new material was very similar in morphology to the original specimens of Tubifex harmani from the USA, but not always in accordance with the text of the original description. The broad ventrolateral folds of the genital segments, not mentioned in the original description, were indeed observed in the holotype during this present study. The total segment number was sometimes higher (up to 125, instead of 95 as noted in the original description), and the number of regenerating anterior segments was more variable (mostly 8, instead of 10) in the original material than discussed by Loden (1979) in his original description. The unusual thickness of the bifid chaetae noted in text by Loden—up to 10 μm in the anterior segments and 9–12 μm in the posterior segments—seems exaggerated when compared to that presented in Fig. 1 View FIGURE 1 D–E in his paper. Indeed, the actual thickness of the bifid chaetae proved to be about 5 μm in posterior, and even less in the anterior segments. The unusual beginning of the clitellum, already in 9/10 (i.e., before the spermathecal pores, according to Loden’s original 1979 description), was not confirmed in the holotype during this present study. The position of the atria in XIII, as noted in the original description, must certainly be a misprint, as only their ends reached XII and XIII on either side in the holotype. The long thin tails of the spermatozeugmata observed in the Dutch material were also seen in the holotype during this present study, but had not recognized or discussed by Loden (1979).
The vas deferens of Tubifex harmani was described by Loden (1979) as being about as long as the atrium but much thinner, with a uniform diameter, and internally ciliated only in its proximal portion. The atrium should have an ejaculatory duct. In Fig. 1 View FIGURE 1 A of Loden (1979), the atrium is depicted as uniformly long and tubular, and with the ejaculatory duct differing from the atrium proper only in its lack of transversal muscular fibres. During the present reexamination of the holotype, the striped appearance of the atrium can also be traced in XI, unlike that in Fig. 1 View FIGURE 1 A of Loden (1979). The male duct may have been drawn by Loden after a dissected specimen—perhaps inteferring this character. The histological study of the new material during this present study did not confirm the bipartite nature of the vas deferens or of the atrium.
Position in the genus
Loden (1979) hesitated between the genera Tubifex Lamarck and Isochaeta Pointner (the latter modified as Isochaetides by Hrabe 1966) to place his new species harmani . Tubifex was chosen owing largely to the uncertain definition of Isochaeta (it is now divided into Isochaetides and Varichaetadrilus ) at that time. After revising the description of this taxon, above, the reproductive system of Tubifex harmani fits well with that in the description of the genus Varichaetadrilus . I suggest to treat this species as Varichaetadrilus harmani n. comb.
This genus was created by Brinkhurst (1981) as Varichaeta ; because that generic name was preoccupied ( Diptera , Tachinidae ), it was renamed Varichaetadrilus by Brinkhurst & Kathman (1983). It was characterized by presence of hair and pectinate chaetae (according to more recent data, however, this is not true for all species), long tubular atria with a small subapically attached prostate gland, distinct penes with sheaths, lack of modified spermathecal chaetae, and lack or only slightly modification of penial chaetae. Prior to this present study, eight valid species had been attributed to the genus:
Varichaetadrilus angustipenis ( Brinkhurst et Cook, 1966) [originally as Limnodrilus
angustipenis in Brinkhurst & Cook (1966), transferred by Brinkhurst (1989)];
V. bizkaiensis Rodriguez et Giani, 1984 ;
V. fulleri Brinkhurst et Kathman, 1983 ;
V. israelis Brinkhurst, 1971 [originally as Isochaeta israelis in Brinkhurst (1971),
transferred by Brinkhurst (1981), and Brinkhurst & Kathman (1983)];
V. minutus ( Brinkhurst, 1965) [originally as Psammoryctides minutus in Brinkhurst
(1965), transferred by Brinkhurst (1987); synonym Isochaeta nevadana Brinkhurst,
1965, according to Brinkhurst (1987)];
V. pacificus (Brinkhurst, 1981) [originally as Varichaeta pacifica in Brinkhurst (1981),
type for the genus, transferred by Brinkhurst & Kathman (1983); including the form
bifidus Brinkhurst et Kathman, 1983];
V. potatiensis (Sokol’skaja, 1983) [originally as Ilyodriloides potatiensis in
Sokol’skaja (1981), used in a new combination without a formal explanation by Timm
& Finogenova (1987)];
V. psammophilus (Loden, 1977) [originally as Limnodrilus psammophilus in Loden
(1977), used in a new combination without a formal explanation by Brinkhurst (1986, 1989)].
V. h a r m a n i (Loden, 1979) n. comb. [originally as Tubifex harmani in Loden (1979)], in
this paper, is the ninth species in this genus.
Six of these species are Nearctic, while three are found in different parts of the Old World: V. bizkaiensis in SW Europe, V. i s r a e l i s in SW Asia, and V. p o t a t i e n s i s in NE Asia.
Six of the nine Varichaetadrilus species ( bizkaiensis , harmani , israelis , minutus , pacificus in its typical form, and potatiensis ) have dorsal hair and pectinate chaetae; however, Brinkhurst & Kathman (1983) assumed that this character can vary intraspecifically, as found in V. pacificus . In V. i s r a e l i s the hair chaetae are serrate. A full set of bifid crotchets at the spermathecal and male pores, like in V. harmani , is noted also in bizkaiensis , minutus , pacificus (although slightly modified in XI, in all three), and possibly also in israelis . Coexistence of the penial sheaths and the ventral chaetae in XI is unusual in Tubificidae .
The flat ventrolateral folds of genital segments, highly characteristic of V. h a r m a n i, have not been described in any other species of Varichaetadrilus . However, they are actually present in V. bizkaiensis (see Fig. 3 View FIGURE 3 by Rodriguez & Giani 1984). They also may have remained unnoticed in some other species (as happened in the original description of V. harmani ). V. bizkaiensis also shares the lateral position of spermathecal pores with V. harmani . In the other species, these pores either lie near the ventral chaetae, or their location is unknown. V. bizkaiensis is the only European congener known so far. It differs from V. harmani in several characters: longer upper tooth in the anteriormost ventral chaetae as well as in those of XI; shorter and thinner vas deferens; distinct anatomical and histological division of the atrium into two regions; short spermathecal duct equipped with an ectal vestibulum; shorter spermatozeugmata without any long tail; and lack of a distinct penial sheath.
The penial sheath is long and thin in V. angustipenis , short and conical in V. minutus , and extremely short in V. israelis . Short sheaths with a narrower midpart, like in V. harmani , are described in the four species: fulleri , pacificus , potatiensis , and psammophilus . Among them, 1) V. f u l l e r i differs from V. harmani , in having only bifid chaetae, spermathecal pores in the place of missing ventral chaetae in X, the presence of a proximal valve in the spermathecal duct, and distally wider vas deferens [apparent in Fig. 1 View FIGURE 1 B of Brinkhurst & Kathman (1983)]; 2) V. potatiensis , in lack of ventral chaetae in X, and in ventrolateral position of the spermathecal pores (Sokol’skaja 1983); 3) V. psammophilus , in having only bifid chaetae (probably lacking ventrally in XI, as the species was ascribed to Limnodrilus by Loden 1977), and long thin vas deferens; and 4) V. pacificus , in more numerous chaetae (including up to seven slightly enlarged ventral, possibly penial chaetae in XI), large penis with a soft sheath folded when constricted, short spermathecal duct with wider vestibulum, and spermatozeugmata of medium length, without a long tail (Brinkhurst & Kathman 1983).
The prostate glands are very large in V. harmani as well as in V. bizkaiensis , contrary to the definition of this genus by Brinkhurst (1981). This suggests that the attribute “small” should be deleted from “prostate gland” in the genus definition. Asexual reproduction with architomy (fragmentation) has not been recorded for any other Varichaetadrilus species, besides V. harmani .
Ecological remarks
The IJsbaantje pond (0.37 ha, depth 1 m) is a nature reserve for endangered plants characteristic of pioneer sites in nutrient poor and poorly buffered waters, such as Apium inundatum, Echinodorus ranunculoides and Lythrum portula . It is fed by iron rich seepage water and also receives agricultural runoff with high phosphate concentration. The pond has been overgrown with reed, Phragmites australis . The accumulated organic sediment and the reeds were removed from the pond between 14.08. and 28.08. 2000 in order to restore pioneer environment for rare plants. This led to a drastic decrease in the concentrations of orthophosphates, total P and Ca, and exposed the underlaying sandy substrate. The pH of the water rose from about 6.5 to 8.3 in one year. The protected plants thrived in the pond again (unpublished data by H. van Kleef). Total oligochaete abundance, dominated by Dero digitata (Müller, 1774) and Lumbriculus variegatus (Müller, 1774) , initially decreased from 4431 to 4 ind. m ², but increased again to about 350 ind. m ² in the following year.
Varichaetadrilus harmani was common in the pond before reclamation, both in May and August 2000. Mature specimens were found on 14.08.2000. Eight of the thirteen individuals exhibited regenerated heads or tails. Two weeks later, after reclamation, a single short, crippled individual was collected, with both ends newly regenerated. In 2001, no individuals were collected in May but healthy immatures were found again in August. The material demonstrates summer maturation and the good regeneration ability of this species, as well as the survival of the population after the reclamation of the pond.
Zoogeographical remarks
Varichaetadrilus harmani is most probably a North American species that has been recently introduced to Europe. It had been not identified earlier on this wellstudied continent. The first confirmed distribution record for V. harmani in Europe, an inland pond, is not directly connected with overseas navigation. However, the species is likely more widely distributed in Europe, yet has not been documented, in all probability because individuals seldom attain sexual maturity (Loden 1979). Even mature individuals can be confused either with Tubifex tubifex (see below) or, owing to their “robust” posterior crotchets, even with one or more Psammoryctides species. I have seen three immature, unidentified tubificid specimens with Tubifex like (but very thick and curved) chaetae and regenerating anterior ends in a sample from the Säveån River at Lerum, Southern Sweden, 10.06.1997 (Erséus et al. 2005). One of them consisted of only eight original segments, with thin regenerated segments at both ends (the anterior end with 10 segments). All three may be V. harmani .
Collado et al. (1999) recorded a PontoCaspian tubificid, Psammoryctides deserticola (Grimm, 1876) , from two North German freshwater lakes not connected with the Ponto Caspian basin. This record is surprising because of the species’ previously known zoogeographical distribution [described by Grimm (1876) from the brackish Caspian Sea, and later known from the Danube basin and the Balkan Peninsula]. It may also be the result of a misidentification of immature (?) tubificids with “robust” posterior crotchets and devoid of modified genital chaetae, as in both V. harmani and the large specimens of Tubifex tubifex described below.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Varichaetadrilus harmani ( Loden, 1979 )
| Timm, Tarmo 2006 |
Tasserkidrilus harmani:
| Brinkhurst 1986 |
V. bizkaiensis
| Rodriguez et Giani 1984 |
V. fulleri
| Brinkhurst et Kathman 1983 |
harmani
| Loden 1979 |
V. israelis
| Brinkhurst 1971 |
Varichaetadrilus angustipenis (
| Brinkhurst et Cook 1966 |
V. minutus (
| Brinkhurst 1965 |