
Acanthodactylus lacrymae, Miralles & Geniez & Beddek & Aranda & Brito & Leblois & Crochet, 2020
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4747.2.4 |
|
publication LSID |
lsid:zoobank.org:pub:F0B392D2-E510-42FA-B252-0ADA001EE43A |
|
DOI |
https://doi.org/10.5281/zenodo.3704097 |
|
persistent identifier |
https://treatment.plazi.org/id/039DEF5B-5F5B-0C19-62AB-6024F737FF77 |
|
treatment provided by |
Plazi |
|
scientific name |
Acanthodactylus lacrymae |
| status |
sp. nov. |
Acanthodactylus lacrymae sp. nov.
(Appendixes I, II; Figs. 6 View FIGURE 6 , 8 View FIGURE 8 , 9 View FIGURE 9 )
Holotype. Muséum national d’Histoire naturelle (Paris), MNHN-RA-2018.0027 (formerly BEV.11991, tissue sample in the BEV tissue collection, code T5986), adult male, collected on May 29th 2012 by Philippe Geniez and Aurélien Miralles ( Fig. 8B View FIGURE 8 ) along the north-eastern shore of the lake of Tislit , north-eastern High Atlas chain, north of Imilchil, Morocco, 32.1986°N, 5.6292°W (WGS84), 2,260 m a.s.l. GoogleMaps
Paratypes. Fifteen individuals, BEV.11980–11990, 11992–11995 (tissue sample codes T5975–5985, 5987– 5990), collected at the type locality at the same date by the same observers.
Additional material. Twelve specimens: BEV.11996–12006 (tissue sample codes T5991–6001) from the western shore of the lake of Izely, Morocco, 32.2182°N, 5.5501°W, 2,276 m a.s.l, collected on May 29 th 2012 by P. Geniez and A. Miralles; photo PGe.1908, live subadult from the type locality GoogleMaps .
Moreover, we identified a number of specimens as A. lacrymae on the basis of morphology, distribution and ecology: BEV.4922–4923 from the Akkal Ouiad valley, north-west of Anefgou, 32.30°N, 5.42°W, 2,200 m a.s.l., collected on April 4 th 1961 by Georges Pasteur; BEV.4924 from Agoudim, south of Tounfite, 32.3763°N, 5.1797°W, 2,100 m a.s.l., collected on April 4 th 1961 by G. Pasteur; BEV.4927–4931 from a few km north of the Tizi n’Talremt, 32.61°N, 4.51°W, no elevation reported, collected on May 28 th 1962 by Jacques Bons; photo PGe.1541 from Jbel Aberdouz, 2,068 m a.s.l., 32.18222°N, 5.20532°W, photographed on July 7 th 2017 by Michel-Ange Bouchet; BEV.4921, from Aït-Moualil, near Bou-Drâa de l’Oudrhès, around 32.53°N, 5.19°W, at least 1,900 m a.s.l. (eastern High Atlas), collected on April 3 rd 1961 by K. Klemmer and G. Pasteur.
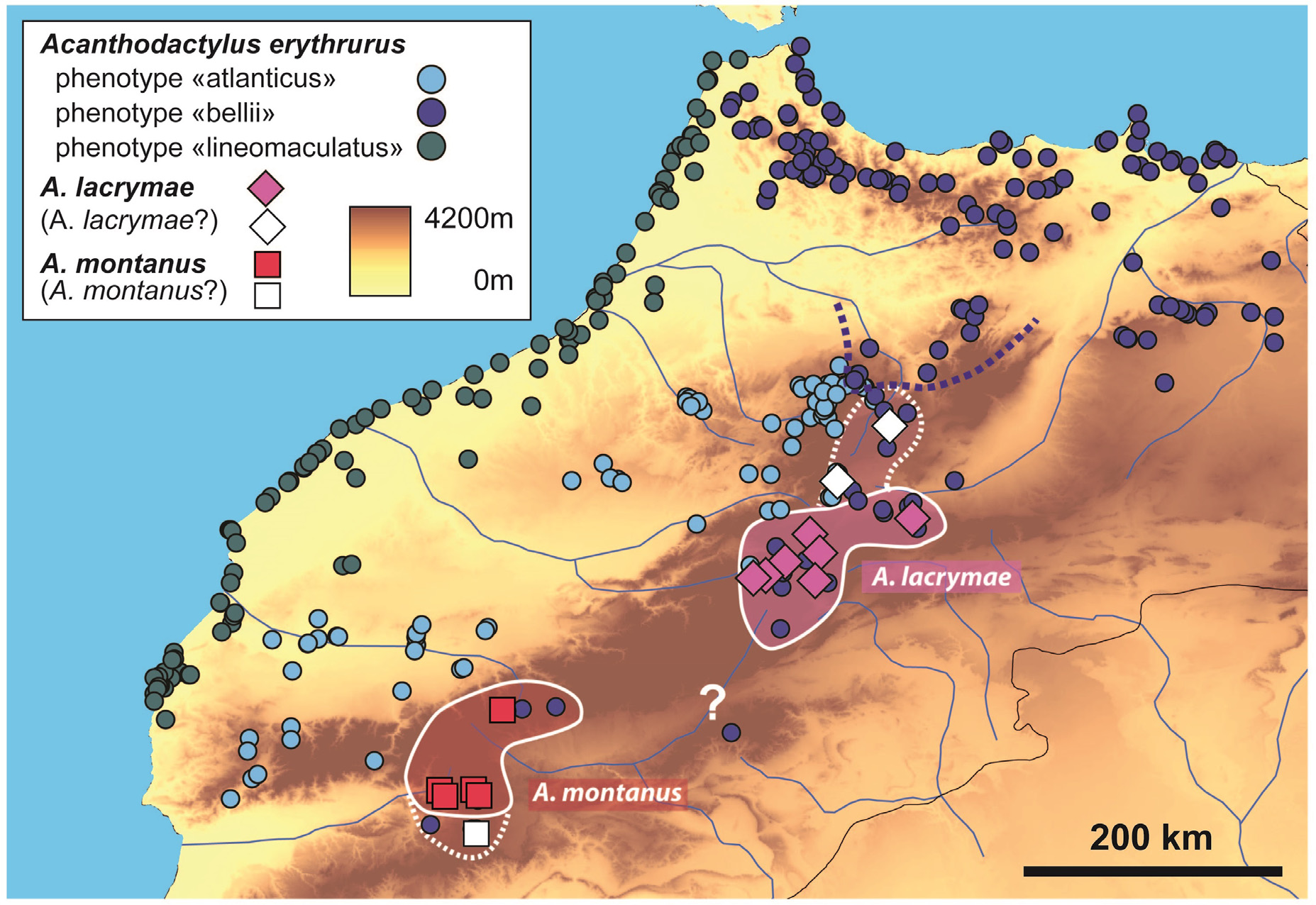
In addition, two series of specimens exhibiting morphological features of A. lacrymae originate from outside the main distribution of the new species: we list them here but the identification is tentative only and they are represented by white diamonds in the map in Figure 10 View FIGURE 10 : BEV.4964–4966 from Itzer (presumably the area around Itzer), approx. 32.88°N, 5.05°W, no elevation reported, collected on June 5 th 1964 by Mr. Cauro; BEV.4920, from 14 km after Boulemane going towards Midelt, 33.25°N, 4.69°W, no elevation reported, collected on April 2 nd 1961 by G. Pasteur.
Etymology. The specific epithet “ lacrymae ”, noun in apposition in the plural form (“tears” in English), comes from a local Berber legend, according to which two young people from different tribes fell in love, but were forbidden to see each other by their families. The grief led them to cry themselves to death, their tears being at the origin of the neighbouring lakes of Tislit (hers) and Isli (his).
Diagnosis. A new species of the Acanthodactylus erythrurus species-group (small flat or carinated dorsal scales; three series of scales around the fingers; three entire supraoculars; 8–10, sometimes 12 straight longitudinal rows of ventrals; slightly pectinate toes; tail and underparts of the hind limbs red or reddish in juveniles, subadults and young adults) from the north-eastern part of the High Atlas Mountains of Morocco above 2,000 m a.s.l., characterized by the combination of the following characters: (1) head scalation always of the “ bellii ” type (subocular in large contact with the upper lip, wedged between the 4 th and 5 th supralabials), one internasal plate, no scales inserted between the prefrontals; (2) a low number of scales, especially for the dorsal scales (44–55 longitudinal rows around the body, mean 50.7), the femoral pores (16–23 or 17–23 on each side, means 19.5 and 19.4) and the subdigital lamellae (19–23, mean 19.8); (3) always less than two rows of supraciliary granules on each side and a reduced number of scales around the supraoculars (15–29 on each side, mean 22.05); (4) smooth temporals; (5) smooth dorsal scales on the neck and back, sometimes slightly but distinctly carinated (tectiform) on the back; (6) stocky proportions, especially in adults, with a relatively short tail (45.6–58.5% of the total length, mean 53.9%); (7) base of the tail extremely enlarged and thick in adult males; (8) in juveniles, undertail coral red turning paler toward the base, even paler on the rear of the underside of the thighs, where it does not reach the rows of femoral pores; undertail white in adults of both sexes; (9) some marginal ventral plates yellow or yellowish in adult and subadult males, sometimes in juveniles (but not clearly so in females); (10) a series of small rounded ocelli on each flank (3–10 brightly coloured scales), usually yellowish in adults, rarely grey greenish, which tend to disappear completely in old specimens.
Body coloration is similar to the other members of the Acanthodactylus erythrurus group. Juveniles are dark (black or blackish) to brown with three pale continuous lines on each side of the body (one on lower flanks, one at the junction between the dorsum and the flanks continuing along the tail, and one on the dorsum joining with the opposite one on the tail base) and pale mid-vertebral area starting on the nape where it is often entirely divided by a dark line and often disappearing on mid-body or before the tail; one or two series of pale ocelli are found in the dark interspaces between these stripes. Adults are medium brown with faint pale lines on the dorsum and upper flanks (corresponding with the juvenile pattern) and a better marked line on the lower flanks; irregular dark marks are present on the back between the pale lines, often leaving the area on the centre of the dorsum unmarked. The flanks typically appear darker due to the large dark areas surrounding each pale ocelli. In many specimens, the pale dorso-lateral lines continue on the outer edge of the pileus to the back of the eye and continue in a faded way along the tail. The pileus is the same colour as the back, usually with dark spots.
The combination of characters listed above separate most specimens from all other members of the A. erythrurus complex (except A. montanus sp. nov. which is extremely similar). Compared to A. montanus , A. lacrymae differs in (1) the parietals and sometimes supraoculars of juveniles with a more continuous and contrasting dark band along their outer edge, creating a more contrasted striped pattern with the pale dorsolateral band continuing through the supratemporals along the sides of the pileus (in A. montanus , parietals have a less distinctly striped pattern of dark marks and light reticulation, see Fig. 9 View FIGURE 9 ); (2) the colour of the base of the undertail is paler and less extensive around the cloacal slit in juveniles (more intense orange red, more similar to the rest of the tail, and more extensive in juveniles A. montanus , Fig. 9 View FIGURE 9 ); (3) the supralabials are unmarked, pale creamy even in juveniles (labials have diffuse dark stripes which persist in adults of A. montanus ); (4) a sharper and better delimited border between the pale lower surface of the tail and a dark band running along the side of the tail, with a complex pattern in males (in A. montanus the pattern is less contrasted, especially in males).
Description of the holotype. An adult male ( Fig. 8B View FIGURE 8 ) measuring 54.0 mm of snout-vent length, 13.8 mm of pileus length, 7.9 mm of pileus width, 7.5 mm of head height, 16.5 mm of forelimb length, 25.6 mm of hindlimb length, with a relatively short non-regenerated tail (63 mm, representing only 54% of the total length), presenting the following scalation features: 54 longitudinal rows of smooth dorsal scales at mid-body, 23 gular scales counted along a line from the contact between the fourth pair of maxillary scales to the collar, 28 transversal rows of ventral plates from the collar to the anal plates, 21 femoral pores on each side, 20 subdigital lamellae beneath the fourth toe, 1 row of supraciliary granules on each side, with 29 (left) and 28 (right) granules, respectively, one entire internasal, no scale inserted between the two prefrontals, two supralabials in contact with the subocular, the latter in large contact with the upper lip on each side. Base of the tail extremely thick. Coloration in life: general ground colour light rufous brown with 6 longitudinal whitish to light grey stripes; the vertebral area between the two central light stripes is nearly uniform with only some vestigial black marks; a series of black transversal marks on the sides of the back; a series of 10 small light greenish ocelli, each surrounded by a large black spot, along the flanks; pileus of the same colour as the dorsum with some black and light marks corresponding approximately to the extension of the dorsal stripes; underparts pure white, a few marginal ventral plates pale yellow or with a black mark; colour delimitation between the side of the body and the underparts very strong and contrasted; a black longitudinal stripe runs along the side of the tail, the top of this stripe slightly wavy on the anterior part and straight on the rest of the side of the tail.
Distribution. Until now, known only with certainty from a handful of localities in a narrow stretch of the eastern High Atlas of Morocco; at high elevations (above 2,000 m a.s.l. for all localities whose elevation is precisely known) between Tislit Lake to the west and the Tizi n’Talremt Lake to the east ( Fig. 10 View FIGURE 10 ). Based on the distribution of animals with the bellii -like subocular conformation in the eastern High Atlas, the potential distribution extends south to the upper Todra Valley (continuous white line in Fig. 10 View FIGURE 10 ). If the identity of the specimens originating from the surroundings of Itzer and the area south of Boulemane (white diamonds in Fig. 10 View FIGURE 10 ) is confirmed by further data, the distribution would extend to the southern edge of the Middle Atlas, potentially in sympatry with the local populations of the IM clade (doted white line in Fig. 10 View FIGURE 10 ).
Natural history. Acanthodactylus lacrymae lives on high plateaus above 2,000 m a.s.l. In both of the visited localities the habitat was similar: open, earthy or gravel ground with short grazed vegetation interspaced with barren patches, numerous stones and low shrubs/patches of taller plants ( Artemisia herba-alba and Hormathophylla spinosa ) that the lizards use as shelters. With its stocky proportions, A. lacrymae runs relatively slowly compared with other members of the A. erythrurus complex, which makes them easier to catch. Both visited populations were dense, with numerous individuals in activity (respectively 17 and 11 specimens caught and others seen but not captured in around 0.01 km ²). These high densities may be explained by the well-preserved habitats typical of high altitudes areas in the region and the absence of direct competitors (the only other observed Lacertid was the North- African wall lizard Podarcis vaucheri , which prefers rocky areas or dense vegetation).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |