
Anaspides spinulae Williams, 1965
|
publication ID |
https://doi.org/ 10.3853/j.2201-4349.68.2016.1669 |
|
persistent identifier |
https://treatment.plazi.org/id/03CDD45B-7076-1C51-FC91-9EFA09E0FCF2 |
|
treatment provided by |
Felipe |
|
scientific name |
Anaspides spinulae Williams, 1965 |
| status |
|
Anaspides spinulae Williams, 1965
Figs 24–28 View Figure 24 View Figure 25 View Figure 26 View Figure 27 View Figure 28 , 36 View Figure 36
Anaspides tasmaniae . — Powell, 1946: 84.
Anaspides spinulae Williams, 1965a: 117–123 , fig. 5 (type locality: Lake St Clair, S of pumping station). — Williams, 1974: 84–85, tab. 4.1. — Knott, 1975: 157 (Lake St Clair specimen only), 173. — Michaelis, 1985: tab. 2. — Richardson, 1985: 3. — O’Brien, 1990: 11–18, tab. 2.2, 2.4, pl. 2. —Jarman & Elliot, 2000: fig. 4, tab. 1. — Camacho et al., 2002: fig. 1, tab. 1. — Lake et al., 2002: 11. — Serov, 2002: 8, 15. — Camacho, 2006: 4.
Type station, 42°06.6'S 146°12.0'E, 3–4.5 m, coll. J.H. Wilson & W.D. Williams, 8 Feb 1963. PARATYPES: AM P14147, 1 juv.♂ (12 mm),type locality GoogleMaps ; AM P14148, 1♀ (slide preparation), type locality ; AM P14149, 1 specimen (slide preparation), type locality ; AM P14150, 1♀ (slide preparation) ; AM P14151, 1 specimen (slide preparation), type locality ; AM P14152, 4 specimens (slide preparation),type locality ; AM P14153, 1♀ (slide preparation), type locality ; AM P14154, 4 specimens (slide preparation), type locality ; AM P14155, 1 juv. ♂ (c. 11 mm), type locality ; AM P14156, 1♀ (21 mm) , 1 juv. ♂ (c. 11 mm), type locality.
Other material examined. YPM 9194 View Materials , 2♂♂ (13–14 mm), 1♀ (15 mm), Lake St Clair , coll. J.H. Wilson, Mar 1961 ; TMAG G124 View Materials , 2 juv. ♂♂ (11–13 mm), 5♀♀ (9–18 mm), Lake St Clair , bottom drag at midnight,coll. A.W.G. Powell, 24 Jul 1937 ; TMAG G134 View Materials , 2 juv. ♂♂ (10–12 mm), Cynthia Bay , Lake St Clair, 42°06.6' S 146°10.1'E, bottom drag, coll. A.W.G. Powell, 26 Aug 1937 GoogleMaps ; AM P99312, 2 juv. ♂♂ (8–9 mm), Lake St Clair , 42°06.75' S 146°11.81' E, 5 m depth, Lake bottom, towed net, 730 m asl, coll. S. Jarman GoogleMaps ; AM P99313, 16 juv. ♂♂ (7–9 mm), 11 juv. ♀♀ (7–10 mm), Lake St Clair , S of Pumphouse Point, 42°06.38' S 146°12.10'E, 2 m, weed bed, on scuba, 730 m asl,coll. M. Driessen & J. Andrew GoogleMaps ; MCZ IZ:68029, 1 juv. ♂ (9 mm), 1 juv. ♀ (8 mm), Lake St Clair , S of Pumphouse Point, 42°06.38' S 146°12.10'E, 2 m, weed bed, on scuba, 730 m asl, coll. M. Driessen & J. Andrew, late 1990s to pre 2005 GoogleMaps ; TMAG G6325 View Materials , 1♂ (23 mm), 1♀ (23 mm), Ida Bay, Lake St Clair , 42°01.7'S 146°08.5' E, 3–7 m, sand & under rocks, stones, plentiful to 7 m, 737 m asl, coll GoogleMaps . R. Holmes , 20 Mar 1990 ; TMAG G6327 View Materials , 1♀ (22 mm), 2 juv. ♀♀ (10–11 mm), Lake St Clair , 42°01.75'S 146°06.94'E, 6 m, between rocks & stones, “No.4”, coll. D. O’Brien, 20 Mar 1990 GoogleMaps ; TMAG G6328 View Materials , 1♂ (14 mm), 1♀ (20 mm), 2 juv. ♀♀ (9–11 mm), Lake St Clair , 42°04.63'S 146°10.02'E,under logs, 6–12 m, coll. D. O’Brien & M. Driessen, 20 Mar 1990 GoogleMaps ; TMAG G6326 View Materials , 1 juv.♂ (11 mm), Lake St Clair , 42°05.08' S 146°11.76'E, 6 m, pebbles and stone outcrops, coll GoogleMaps . R. Holmes , 20 Mar 1990 ; TMAG G6324 View Materials , 3 juv. ♂♂ (9–10 mm), 5 juv. ♀♀ (8–9 mm), Lake St Clair , 42°06.70'S 146°11.74'E, 3–4 m, amongst Isoetes , under stones, rocks, site 9, coll. D. O’Brien, 20 Mar 1990 GoogleMaps ; TMAG G6323 View Materials , 2 juv. ♂♂ (10–11 mm), 2 juv. ♀♀ (11–12 mm), Lake St Clair , 42°01.04'S 146°06.74'E, 4–6 m, stones and pebbles,coll GoogleMaps . R. Holmes , 20 Mar 1990 ; TMAG G6329 View Materials , 1 juv. ♂ (8 mm), 1♀ (20 mm), no locality data .
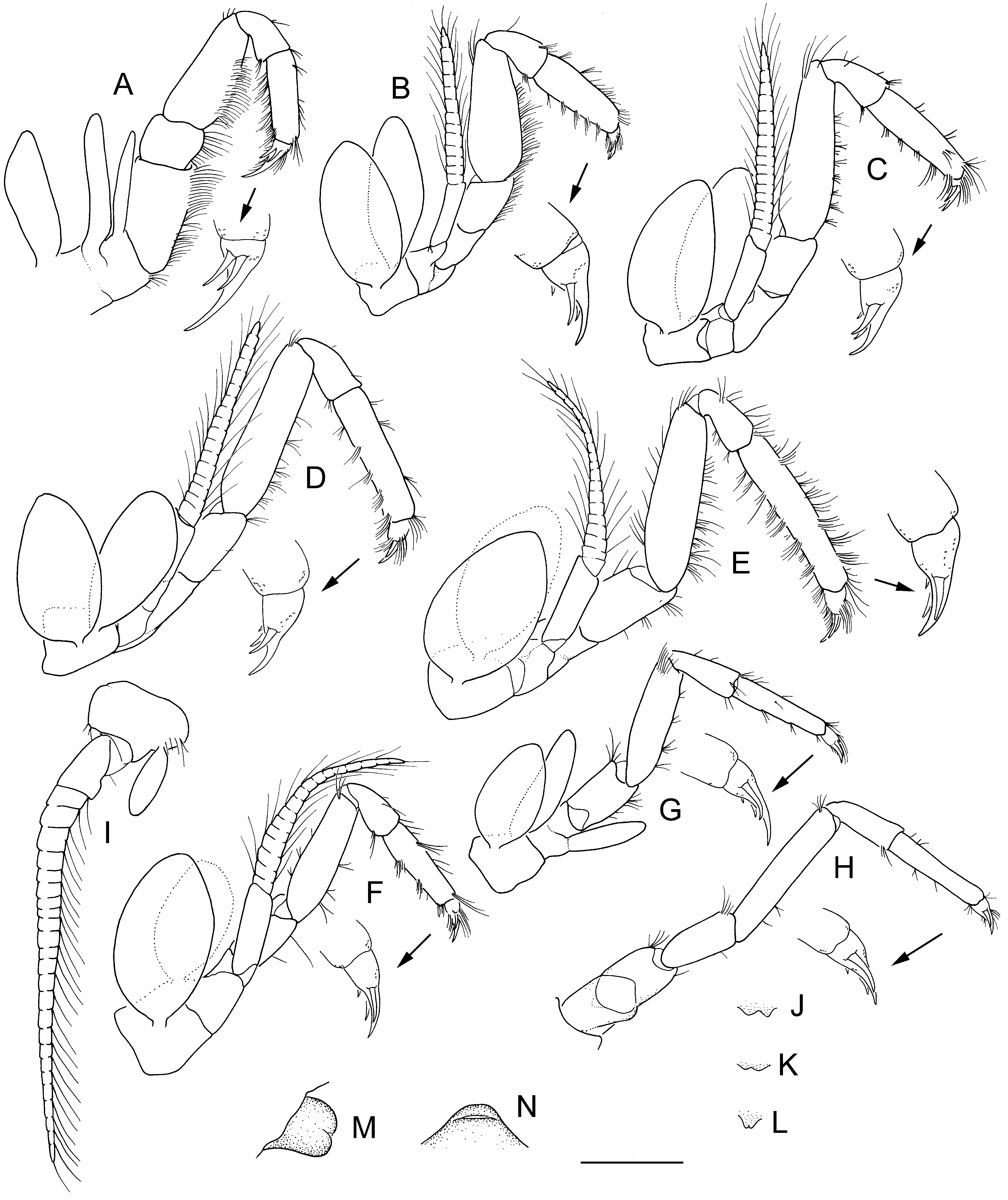
Description. Eyes with large well-developed cornea, pigmented, distinctly wider than stalk, subglobular, longer than half length of stalk; stalk with divergent margins. Rostrum narrow in adults, apex blunt.
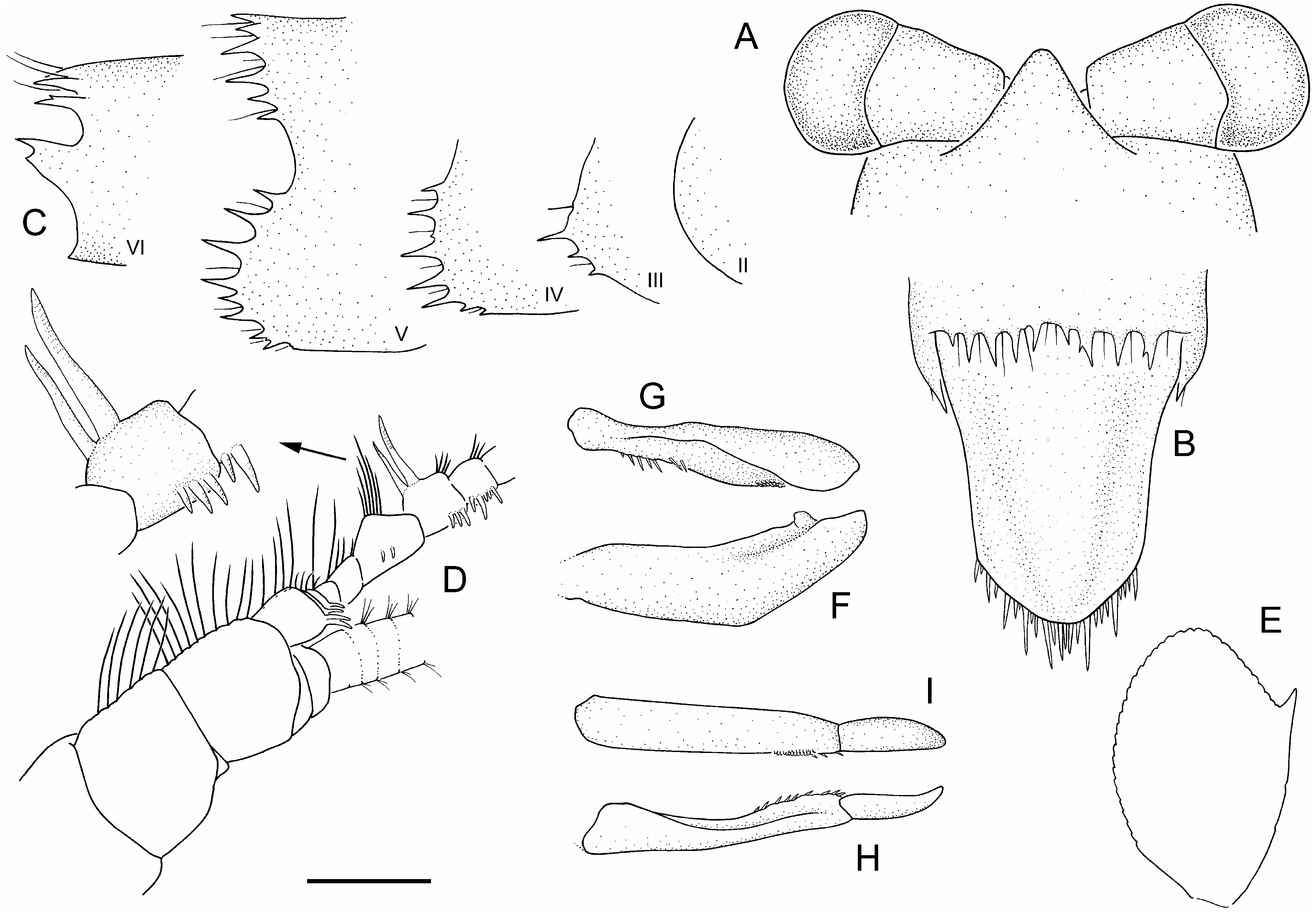
Pleonites with sparsely setose pleural margins, rounded; pleuron 1–2 with 0–3 small spines; pleura 3–5 prominently spinose. Pleonites 5–6 posterior tergal margins prominently spinose, setose. Pleonite 6 posterolateral margin produced to prominent spine, occasionally with secondary spine. Pleonal sternites 3–5 with low, median processes between pleopod bases, distinctly bilobed and widest on sternite 3, bilobed on sternite 4, weakly bilobed and narrowest on sternite 5.
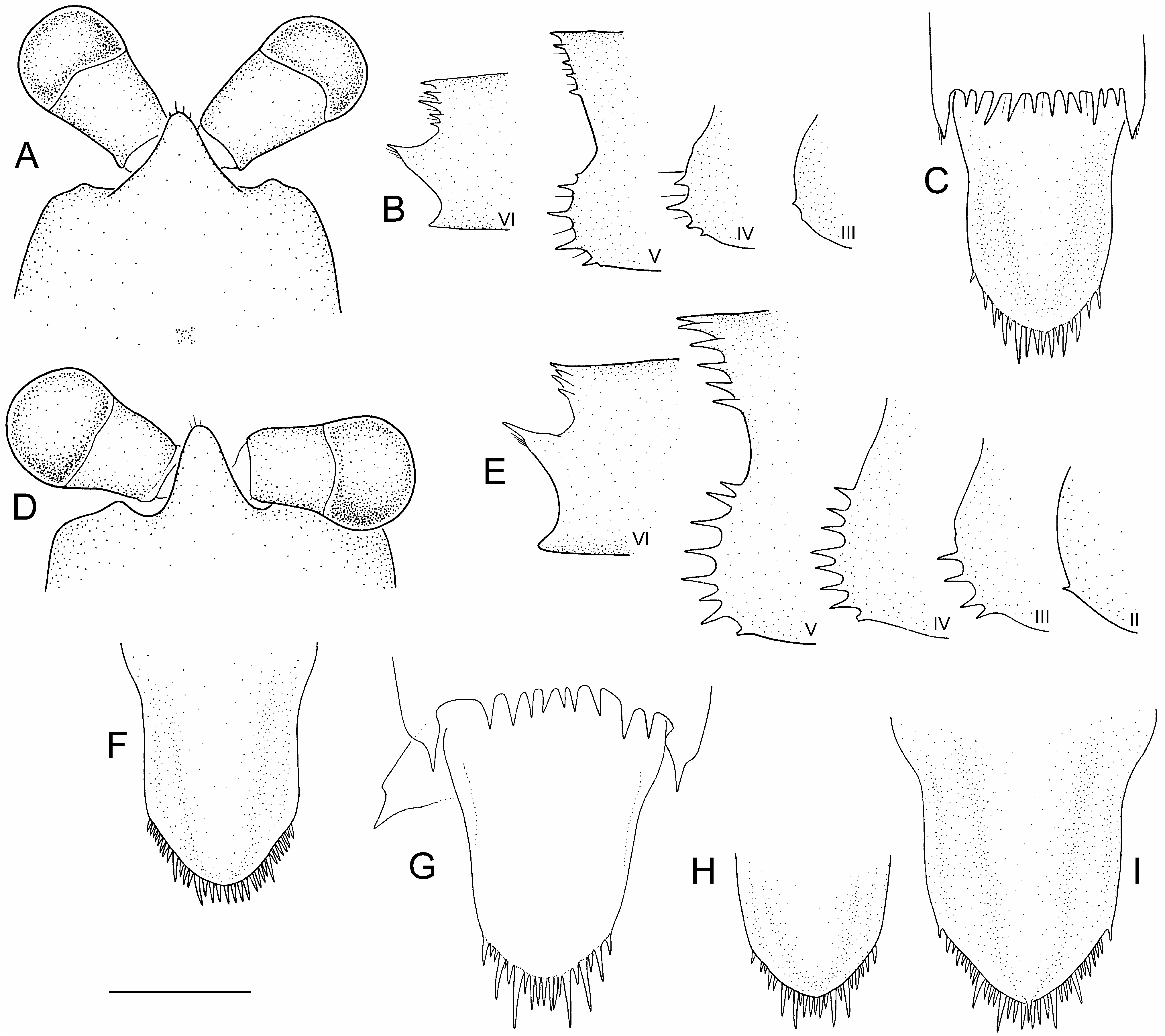
Telson longer than wide, widest proximally; lateral margins sinuous in dorsal outline, distally convergent; transition from lateral to posterior margin obtusely angular; posterior margin angular to slightly rounded, blunt medially; posterior spine row with 17–32 slender, uneven, closely spaced spines, with several longer spines, approximately evenly spaced among shorter spines; rarely with dorsomedian spine above posterior margin.
Antennule inner flagellum about 0.2 × body length (22 articles in holotype); article 7 inner margin obtusely angled in adult males, with 2 long, slender clasping spines proximally; outer flagellum 0.5–0.6 × body length (67–71 articles in holotype). Antennal flagellum 0.3–0.5 × body length (49–50 articles in holotype); scaphocerite elongate, ovate, lateral spine slightly distal to midlength; apex almost reaching or slightly overreaching apex of distal peduncular article.
Right mandibular incisor process with proximal tooth distally bifid or trifid.
Pleopods 1–5 with endopod in adults.Adult male pleopod 1 distally widened, scoop-like, lateral margins weakly expanded, not obscuring retinacular lobe in lateral view.
Uropodal protopod dorsally with 1 or 2 small spines; exopod with 2–4 movable spines on outer margin near position of partial diaeresis; exopod length about 3 times width, as wide as endopod, apex rounded, relatively narrow.
Measurements. Male (n = 35) 7–23 mm, female (n = 36) 7–25 mm.
Remarks. Williams (1965a) distinguished A. spinulae from A. tasmaniae on the basis of its pronounced pleonal spination, particularly of pleura 3–5 and tergites 5–6, uneven telson spine row, and apparently unusual ecology—the lacustrine habitat of Lake St Clair. Anaspides tasmaniae (now known to comprise several species) was thought, at the time Williams wrote, to occupy only surface creeks, tarns and streams, but subsequent discoveries from lakes throughout Tasmania refuted that assumption ( Williams, 1974). The validity of A. spinulae has been questioned ever since ( Williams, 1974; O’Brien, 1990; Jarman & Elliott, 2000). In particular, the observation of a possible “gradation of increasing spination in a southerly direction across the western portion of the Great Western Tiers” down to Lake St Clair cast doubt on the validity of A. spinulae ( Knott, 1975; O’Brien, 1990). Moreover, Anaspides throughout much of Tasmania show some degree of pleural and tergal spination, including A. tasmaniae from Mt Wellington, further eroding the distinctiveness of A. spinulae (see O’Brien, 1990). Others have accepted a limited range for A. spinulae , from Lake St Clair to Clarence Lagoon ( Richardson, 1985; Swain, 2000). The status of A. spinulae has remained debated, partly owing to the seemingly unreliable diagnostic features identified by Williams (1965a), and partly owing to the inadequate and perhaps misleading original account of the species, in which the holotype was not figured.
Review of all available material from Lake St Clair, as well as western Central Plateau specimens reported by Knott (1975) and O’Brien (1990) as A. “ spinulae ” (= A. richardsoni sp. nov.; Fig. 22 View Figure 22 ), indicates that A. spinulae is a valid species. The present concept of A. spinulae emphasizes the male secondary sexual characters in addition to the pleonal spination. Thus, A. spinulae can be distinguished from all other species of the genus by the combination of two antennular clasping spines in adult males ( Fig. 27D View Figure 27 ), prominently spinose pleural margins of pleonites 2–5 and posterior tergal margins of pleonites 5–6, the prominently spiniform posterolateral angle of pleonite 6 and angular posterior margin of the telson ( Figs 25 View Figure 25 , 27 View Figure 27 , 28 View Figure 28 ). The cornea of A. spinulae is also proportionally more inflated than any of its congeners at a similar size, being noticeably wider than the stalk ( Figs 25A View Figure 25 , 27A View Figure 27 , 28A,D View Figure 28 ).
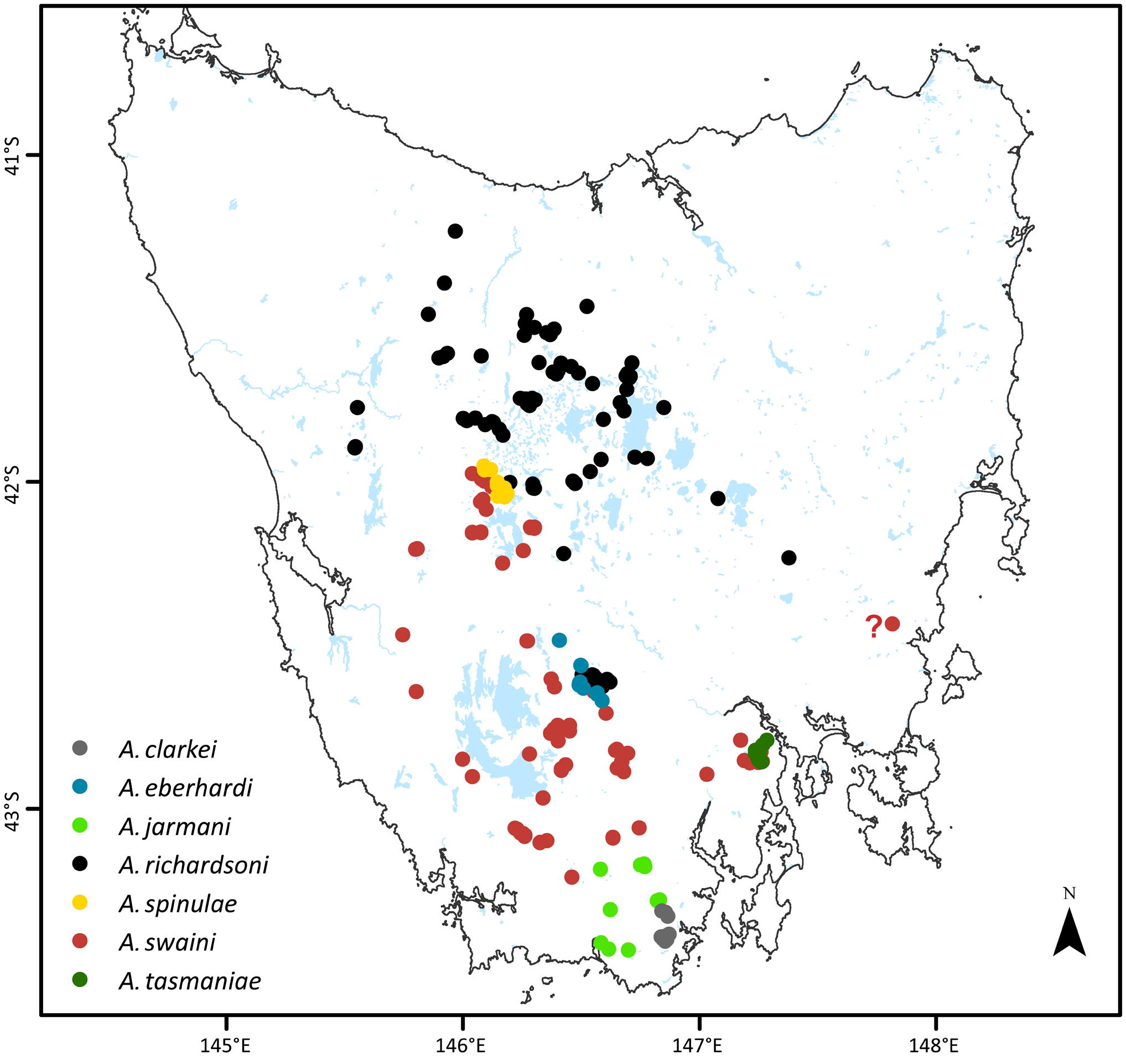
Despite A. spinulae being a valid species clearly separated from A. tasmaniae , its taxonomic boundaries remain to be fully circumscribed.The secondary sexual modifications of the male antennule and presence of the pleopod 5 endopod suggest a close relationship to the northern form of A. swaini occurring to the immediate west and south of Lake St Clair. Although Williams (1965a) hypothesized that A. spinulae survived the Pleistocene glaciation of Lake St Clair in adjacent periglacial lakes or melt-waters, areas currently occupied by A. swaini , A. spinulae might equally have persisted in-situ in deeper parts of the lake during that time. This more easily accounts for the very limited range of the species today, nested between the ranges of A. swaini and A. richardsoni . Knott (1975) alternatively hypothesized A. spinulae to be only a temporary resident in Lake St Clair, being periodically flushed into the lake from adjacent creeks and streams, with its primary habitat being the western Central Plateau. Present results, however, indicate otherwise. The nearest neighbouring Anaspides populations to the immediate south and west ( A. swaini ) and immediate east ( A. richardsoni , eastern side of the Travellers Rest Range) of Lake St Clair have “normal”, non-spiny pleonal ornamentation. The strongly spinose forms (A. “ spinulae ” of O’Brien, 1990) occur further afield on the western Central Plateau to the east and northeast of Lake St Clair, and although closely resembling A. spinulae , are referable to A. richardsoni having one instead of two male antennular clasping spines. It is enigmatic that the strongly spinose “spinulae” morphology is known only in Anaspides from Lake St Clair and the western Central Plateau, whereas the nearest neighbours to the lake all have “normal” pleonal ornamentation. As noted under the account of A. richardsoni , the occurrence of the spiny body form does not have an immediate ecological basis, relating neither to a lacustrine habitat nor presence of fish predators. The observed morphological patterns in Anaspides from west to east of Lake St Clair are presently difficult to interpret, and the limited currently available molecular data are equivocal (Jarman & Elliott, 2000; Andrew, 2005). Thus, the relationship of A. spinulae to neighbouring Anaspides populations, currently assigned to A. swaini and A. richardsoni , respectively, requires more detailed analysis beyond the scope of the present study. Although conceivably more wide ranging, A. spinulae is presently known with certainty only from Lake St Clair.
Previous records of A. spinulae from Clarence Lagoon and elsewhere on the Central Plateau are referable to A. richardsoni . Although resembling A. spinulae in pleonal spination, adult males from Clarence Lagoon, like other spiny Central Plateau forms have a single antennular clasping spine, diagnostic of A. richardsoni . It is notable though that the corneas of spiny A. richardsoni from Clarence Lagoon ( Fig. 22F View Figure 22 ), like true A. spinulae , are proportionally larger and more expanded than size-matched A. richardsoni from elsewhere on the Central Plateau (including a non-spiny specimen from Clarence Lagoon; Fig. 22A View Figure 22 ).
Anaspides spinulae apparently matures at a smaller size than congeners. Development of antennular modifications begins in juvenile males at 11–13 mm, with clasping spines appearing sequentially and with increasing curvature of the proximal portion of the flagellum. By 14–15 mm body length, the male secondary sexual characters of A. spinulae are well-developed suggesting sexual maturity. Males of other species of Anaspides do not attain similar development until 18 mm or larger. Possibly also significant is that the largest known specimen of A. spinulae , at 25 mm, is considerably smaller than the largest specimens of congeners (35 mm, A. tasmaniae ; 38 mm, A. clarkei ; 47 mm, A. eberhardi ; 31 mm, A. jarmani ; 55 mm, A. richardsoni ; 40 mm, A. swaini ).
Specimens of A. spinulae are in most respects morphologically uniform apart from typical allometric variation in the slenderness of the rostrum (increasingly slender with increasing size), eye size (proportionally larger in smaller specimens), and pleonal spine length (proportionally longest in the smaller specimens). The endopod of pleopod 5 is absent or rudimentary in juveniles up to 10 mm body length; present in all others. Pleonal spination is pronounced, even in the smallest juveniles, with pleural spines always present on pleonites 3–5, and those on pleonite 2 appearing at about 20 mm body length. The most significant allometric changes are in the spination of the posterior margin of the telson. The telson spination in the smallest specimens is markedly uneven, with 4–6 long spines evenly distributed among the shorter remaining spines ( Figs 27B View Figure 27 , 28C,G,H View Figure 28 ). With increasing body size, the longer telson spines become shorter and more similar to surrounding spines, as in the holotype, somewhat approaching adults of other epigean species ( Figs 25B View Figure 25 , 28F,I View Figure 28 ). Similar, albeit less marked changes, are also evident in other species, such as A. richardsoni and A. swaini , indicating a general developmental pattern. Apart from allometric changes, A. spinulae is morphologically uniform, with few other observed variations. The posterolateral spine on pleonite 6 may be accompanied by a secondary spine, and the uropodal protopod and the outer margin of the uropodal exopod have 1 or 2 and 2–4 spines, respectively.
Unfortunately, Williams’ original characterization of A. spinulae as having a strongly uneven spine row on the telson was misleading, because his accompanying unscaled illustration ( Williams, 1965a: fig. 5E) was of a small dissected juvenile (estimated c. 10 mm body length; AM P14152; Fig. 28G View Figure 28 ) in which the differences in spine length are most pronounced. At any given size, the posterior telson spines in A. spinulae are less regular in length than in other epigean Anaspides of similar size, but not nearly as pronounced as would be assumed from Williams’ figure. Subsequent reports (e.g., O’Brien, 1990) of telsons “intermediate” between A. spinulae and typical A. tasmaniae were based on the implication from Williams’ (1965a) figure that an adult was depicted. Further, Williams’ (1965a) description of the pleonal setae of both A. tasmaniae and A. spinulae as spines further clouded distinguishing the two forms. This possibly contributed to some of O’Brien’s (1990) difficulties in distinguishing between what he called A. spinulae , A. “ spinulae ” and A. tasmaniae leading to the strong suggestion that all represent the same species. O’Brien (1990) also reported Central Plateau specimens with “longer setae… ”, including specimens from Butlers Gorge. As already noted, however, the length of pleonal setae varies allometrically in Anaspides and is shortest in adults, as corroborated in specimens from Butlers Gorge examined herein (referrable to A. swaini ). Whereas the posterior telson ornamentation of adult A. spinulae does differ from most other congeners in length and regularity, it is not nearly as marked as originally implied by Williams (1965a).
O’Brien (1990) recorded A. spinulae from around the margins of Lake St Clair at about 1.5–15 m depth on multiple substrate types. It apparently does not occur on silted substrates, but is common on stony or pebbled outcrops and among exfoliating sheets of rock on weathering dolerite boulders, with highest densities under rocks, fallen logs and branches, or in Isoetes algal beds.
Distribution. Presently known only from Lake St Clair; 1.5–15 m depth; 737 m asl.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Anaspides spinulae Williams, 1965
| Ahyong, Shane T. 2016 |
Anaspides spinulae
| Camacho, A 2006: 4 |
| Lake, P 2002: 11 |
| Serov, P 2002: 8 |
| O'Brien, D 1990: 11 |
| Richardson, A 1985: 3 |
| Knott, B 1975: 157 |
| Williams, W 1974: 84 |
| Williams, W 1965: 123 |
Anaspides tasmaniae
| Powell, A 1946: 84 |