
Columbella Bellardii, M. Hörnes, 1852
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5295.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:F3A52660-70B8-439F-A7A0-F45ADC975EA5 |
|
DOI |
https://doi.org/10.5281/zenodo.7976057 |
|
persistent identifier |
https://treatment.plazi.org/id/03CF879C-2C71-0922-FF1D-F8CAFDCB768F |
|
treatment provided by |
Plazi |
|
scientific name |
Columbella Bellardii |
| status |
|
Columbella Bellardii View in CoL ? Hoernes— Mayer 1864: 76.
Discussion. According to his discussion, Mayer (1864: 76) had a fragment of a columbellid that he tentatively identified as Columbella bellardii Ĥrnes, 1852, which was assigned as the type species of the genus Auingeria Harzhauser & Landau (2021) . That genus is endemic to the Paratethys, and Mayer’s determination is unlikely. We exclude this record from the assemblage.
Palaeoclimatology
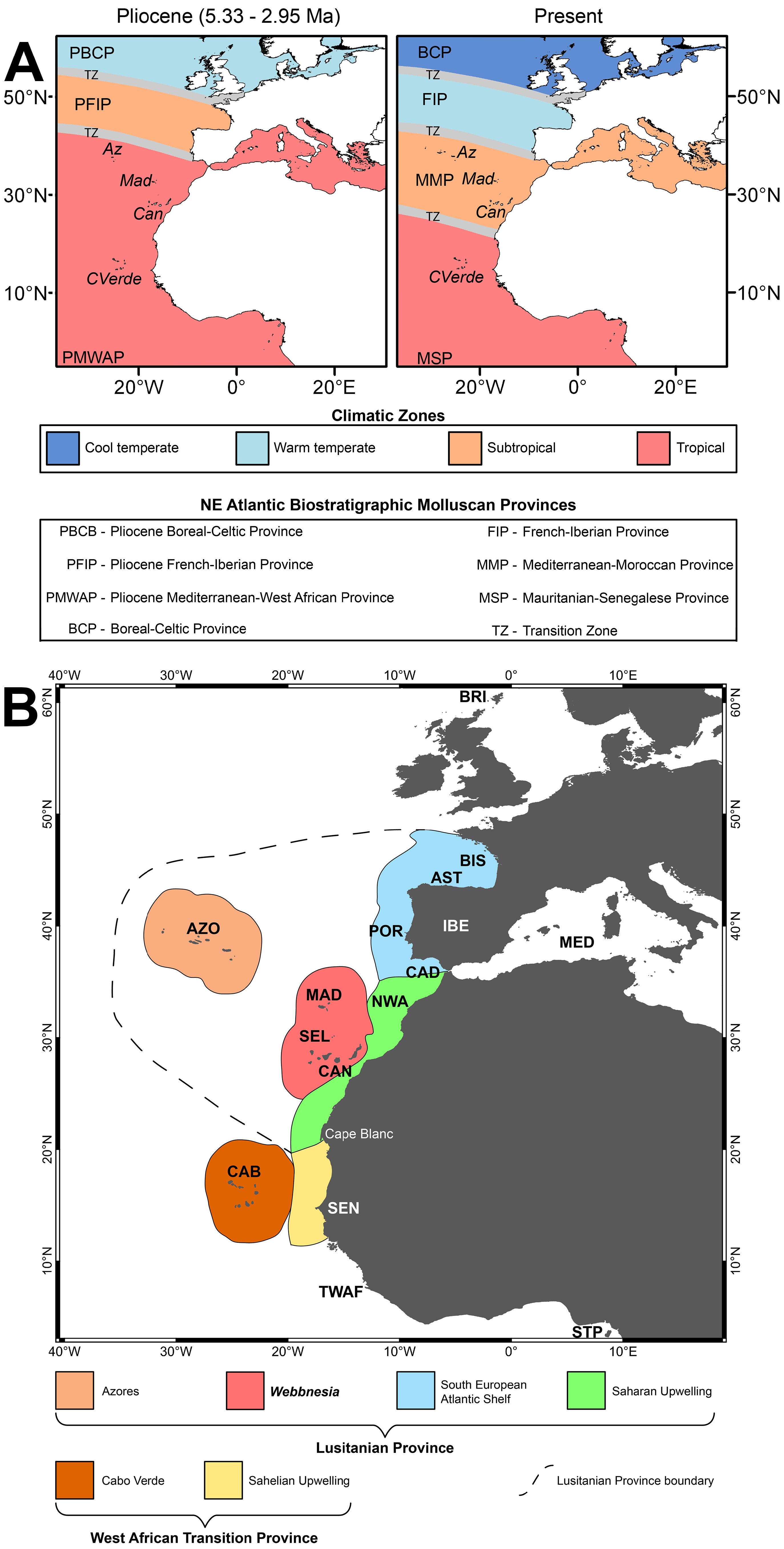
Based on the presence of gastropod genera requiring fully tropical conditions, such as Turbo Linnaeus, 1758 , Nerita Linnaeus, 1758 , Cheilea Modeer, 1793 , Hipponix Defrance, 1819 , Thetystrombus Dekkers, 2008 , Distorsio R̂ding, 1798, Bellardithala Harzhauser & Landau, 2021 , Cancilla Swainson, 1840 , Tomellana Wenz, 1943 , high diversity of conids, including fully tropical genera such as Africonus Petuch, 1975 , Plagioconus Tucker & Tenorio, 2009 and Stephanoconus M̂rch, 1852 and the species Aurantilaria tarbelliana ( Grateloup, 1845) ( Tucker & Tenorio 2009; Landau et al. 2011, table 1; Ávila et al. 2016b; Harzhauser & Landau 2016), We interpret the assemblage as being fully tropical, rather than subtropical as done by Mayer (1864). Some of the species present in the Santa Maria assemblages [e.g., Cheilea equestris ( Linnaeus, 1758) , Hipponix sulcatus ( Borson, 1820) , and Tethystrombus coronatus (Defrance, 1827) ] are present in the Mediterranean during the fully tropical Pliocene but did not survive the cooling event at the end of MPPMU1. Conversely, there are no genera present typical of European temperate waters (e.g., Buccinum Linnaeus, 1758 , Colus R̂ding, 1798, Neptunea R̂ding, 1798).
Palaeo-SSTs for water masses around Santa Maria were discussed by Ávila et al. (2016b) based on the presence of Persististrombus coronatus (Defrance, 1827) , now placed in the genus Tethystrombus Dekkers, 2008. Those authors suggested that during MPPMU1 (i.e., Zanclean and earliest Piacenzian from 5.33 to about 3 Ma) there was a large climatic zone (see Fig. 5A View FIGURE 5 ) with full tropical conditions that extended some distance north of the Azores south to Angola: the Pliocene Mediterranean-West African (palaeobiogeographic) Province (PMWAP) (see also Monegatti & Raffi, 2001; Silva & Landau, 2007). They also stated that the palaeoclimate at Santa Maria Island was drastically different from that seen at those latitudes today, with mean annual sea surface temperatures (SSTs) about 3.7 °C to 6.3 °C higher than the present-day 20.6 °C, and with mean monthly SSTs ranging from 20 °C to 28 °C, with six months with mean SSTs over 24 °C, conditions typical of a tropical setting. The rest of the warm-water taxa represented in the same Santa Maria assemblages further support this conclusion.
Palaeobiogeography
Palaeobiogeographical implications are difficult to draw based on such a small number of taxa. However, certain inferences can be made (cf. Fig. 5A View FIGURE 5 ). The Azorean PMWAP assemblage shows a high affinity with that of the tropical Mediterranean-West African Palaeobiogeographical Province during the Zanclean and earliest Piacenzian (the period of time represented by the MPPMU1). Twenty-seven (35%) of species are found both in the Santa Maria assemblages and in the Pliocene Mediterranean during MPPMU1. The association with the subtropical Pliocene French-Iberian Province (PFIP sensu Ávila et al. 2016b) is weaker [13 species (17%)], and very few species are found in common with the warm temperate Pliocene Boreal-Celtic Province (PBCP sensu Ávila et al. 2016b) further to the North [6 or 7 species (8-9%)], mainly ubiquitous European species such as Calyptraea chinensis ( Linnaeus, 1758) , Petaloconchus intortus ( Lamarck, 1818) , Thylacodes arenarius ( Linnaeus, 1758) , and Tenagodus (Tenagodus) obtusus ( Schumacher, 1817) . We note that at least some of these widely distributed species may represent species groups that are difficult if not impossible to separate based on shell characters rather than single species (e.g., C. chinensis ).
The most widely dispersed species found in the Santa Maria assemblages are all within the Tonnoidea Suter, 1913 . This is hardly surprising as tonnoidean species have large multispiral protoconchs ( Beu 1988: 75) suggesting planktotrophic dispersal, and the veligers of modern species have been shown to live for up to in the plankton—teleplanic larvae ( Jablonski & Lutz 1980), and to travel vast distances ( Beu 1988: 73). Two species have an amphiatlantic distribution during the Plio-Pleistocene: Aspa marginata ( Gmelin, 1791) , which today no longer occurs in the tropical western Atlantic and is found only along the coast of West Africa, Madeira, Canary Islands and Cabo Verde archipelagos; and Monoplex comptus (A. Adams, 1855) which still has an amphiatlantic distribution. Of particular interest is the presence of Distorsio mcgintyi Emerson & Puffer, 1953 in the Azores assemblages. This species has a long history in the Cainozoic of the Tropical American Atlantic, going back to the lower Miocene ( Landau & Silva 2010; Beu 2010). Its presence in the Pliocene of the Azores Archipelago is the most eastern record for the species.
We consider the following to be endemic species to the Santa Maria Pliocene to present-day assemblages: Danilia pterostomus (Bronn, 1861) , Turbo hartungi Bronn 1861 , Thericium crenulosum (Bronn, 1862) , Thericium hartungi (Mayer, 1862) , Thericium incultum (Mayer, 1862) , Bittium miradouroense nov. sp., Bittium an nov. sp.?, Bittium nanum ( Mayer, 1864) , Janthina krejcii Beu, 2017 , Euspira atlantica (Mayer, 1862) , Alvania mediolittoralis Gofas, 1989 , Alvania sleursi ( Amati, 1987) , Erato mayeri nov. sp., Gibberula vignali Dautzenberg & Fischer, 1896 , Cancellaria parcestriata Bronn, 1862 , Fusinus sp. ?, Tritia atlantica (Mayer, 1862) , Tritia aff. striatula striatula ( Eichwald, 1829) , Tritia vetulum (Mayer, 1862) , Cancilla volvaria (Bronn, 1862) , Africonus trochilus ( Mayer, 1864) , and Stephanoconus candidatus (Mayer, 1862) . Taxa considered species inquirenda have been excluded. Pseudopusula parcicosta (Bronn, 1862) has a slightly wider distribution, described from the lower Pliocene of Santa Maria and the Canary Islands ( Fehse & Grego 2014). Therefore, 22 Pliocene gastropod species (29%) are considered as endemic to the Azores, and this compares with a total of 23 Last Interglacial (MIS 5e) endemic gastropod species (19%; Ávila et al., 2015a) and 41 (14.4%) recent endemic gastropod species ( Cordeiro & Ávila, 2015). At genus level, the composition is typical of that of the tropical Pliocene Mediterranean-West African palaeobiogeographic Province, except for the genera Janthina R̂ding, 1798 and Pseudopusula Fehse & Grego, 2014 . However, at species level the assemblage is relatively endemic, with 22 (29%) of the species restricted to the Azores archipelago.
Working on extant faunas, Spalding et al. (2007) devised a global model for coastal and shelf areas constructed of realms, provinces and ecoregions that was refined for the Macaronesian archipelagos by Freitas et al. (2019; cf. Fig. 5B View FIGURE 5 ). Spalding’s use of the term ‘ecoregion’ equates roughly to the ‘subprovinces’ used by palaeobiogeographers (e.g., Woodring 1974; Petuch 1982, 1988, 2004; Landau et al. 2008, 2019).
In the present-day faunas, Freitas et al. (2019: fig. 7) increased the resolution of Spalding et al. (2007: box 1, fig. 3) and recognized the Azores ecoregion and the Webbnesia ecoregion (including Madeira, Selvagens and Canaries) all belonging to the Lusitanian biogeographic Province, with a distinct Cabo Verde biogeographic subprovince included in the West-African Transition biogeographic Province. The Azores ecoregion, the Webbnesia ecoregion the Saharan upwelling ecoregion and the Mediterranean (cf. Fig 5B View FIGURE 5 ), altogether, are roughly equivalent to the subtropical Mediterranean-Moroccan Province of Raffi et al. (1985) (cf. Fig. 5A View FIGURE 5 ), whereas the South European Atlantic Shelf ecoregion (cf. Fig. 5B View FIGURE 5 ) equals to the warm temperate French-Iberian Province (cf. Fig. 5A View FIGURE 5 ). Finally, Cabo Verde subprovince and the Sahelian Upwelling ecoregion ( Fig. 5B View FIGURE 5 ) correspond to the Mauritanian-Senegalese Province ( Fig. 5A View FIGURE 5 ).
This revision of the Santa Maria Pliocene gastropod assemblages further strengthens the hypothesis that the location of the boundaries between distinct biogeographic units, especially of those located along the eastern Atlantic façade (see Fig. 5A View FIGURE 5 ) were already in place at least since the early Pliocene.
At the end of the Pliocene, the tropical Pliocene Mediterranean-West African Province split into the present-day northern subtropical Mediterranean-Moroccan Province (MMP) and southern Mauritanian-Senegalese Province (MSP; see Monegatti & Raffi 2007, 2010; Silva & Landau 2007; Landau et al. 2011; Ávila et al., 2016b). The stability of the biogeographical subprovince units over time, despite the fracture or change in the larger biogeographic unit in which they occur, has also been demonstrated in other areas (e.g., Landau et al. 2008; 2019). The location of the transition zones (i.e., the location of the boundaries between different, consecutive biogeographic areas) along the eastern Atlantic latitudinally also seem to show long-term stability. For example, the transition zone between the Pliocene Mediterranean-West African Province (to the south) and the Pliocene French-Iberian Province (to the north) was located in an area that extended from Lisbon south to Cabo de S„o Vicente (Algarve) (see Fig. 5A View FIGURE 5 ) (e.g., Silva 2001; Monegatti & Raffi 2007; Silva & Landau 2007). The present transition zone between the subtropical Mediterranean-Moroccan Province (at south) and the warm temperate French-Iberian Province (at north) is located precisely along the same coastal area ( Raffi et al. 1985). A similar situation happens for the location of the transition zone between the subtropical Pliocene French-Iberian Province (at south) and the warm temperate Pliocene Boreal-Celtic Province (at north), which is the same as that of the present transition zone between the warm temperate French-Iberian Province (at south) and the cool temperate Boreal-Celtic Province (at north).
The Santa Maria Pliocene assemblages are unique amongst European Pliocene assemblages in containing fossils of the holoplanktonic gastropod genus Janthina , represented by two species: J. krejcii Beu, 2017 and Janthina typica (Bronn, 1861) . Based on the presence/absence of Janthina species Beu (2017) constructed a circumglobal time range zonation, especially useful in the Antipodean area and Japan. Unfortunately, this model is of limited use in the European and Caribbean assemblages where Janthina species do not occur. Nevertheless, the Santa Maria assemblages were crucial in the construction of this zonation, and we note that the end of the J. typica zone (of Beu 2017) roughly coincides with the end of MPPMU1 at about 3 Ma ( Beu 2017: figs. 22, 23) and the end of fully tropical conditions within the Mediterranean and adjacent Atlantic ( Raffi & Monegatti 1993; Monegatti & Raffi, 2001; Silva & Landau 2007; Ávila et al. 2016b).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Columbella Bellardii
| Sacchetti, Claudia, Landau, Bernard & Ávila, Sérgio P. 2023 |
Columbella Bellardii
| Mayer, K. 1864: 76 |