
Erenna insidiator, Pugh, P. R. & Haddock, S. H. D., 2016
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4189.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:0F0EE4C3-05B9-4248-944D-B41DC8361CB2 |
|
DOI |
https://doi.org/10.5281/zenodo.6067132 |
|
persistent identifier |
https://treatment.plazi.org/id/D277A579-4C36-295D-FF68-8FABF1DEE310 |
|
treatment provided by |
Plazi |
|
scientific name |
Erenna insidiator |
| status |
sp. nov. |
Erenna insidiator View in CoL sp. nov.
Diagnosis. Nectophores basic erennid ridge pattern. The upper laterals do not divide nor extend as far as the ostium. Two types of bract, both with pair of lateral cusps in proximal half; larger ones with rectangular distal ectodermal process on upper side, containing nematocysts. Very characteristic tentilla, consisting of a long pedicle and a long tube mostly filled with spongy tissue through which passes the endodermal canal. The tube tapered distally and there enclosing a photophore. Cnidoband itself forms the corrugated outer perimeter on a distinct process from the outside of the distal part of the cylinder.
Material examined. Only two colonies of Erenna insidiator sp. nov. are known to have been collected and their station data are given here.
Part of the specimen from Tiburon Dive 1040 is designated the holotype and will be donated to National Museum of Natural History , Smithsonian Institution, Washington, D.C., U.S.A., while the remainder, and the other specimen will be maintained, in the siphonophore collections at Brown University, Providence, R.I.
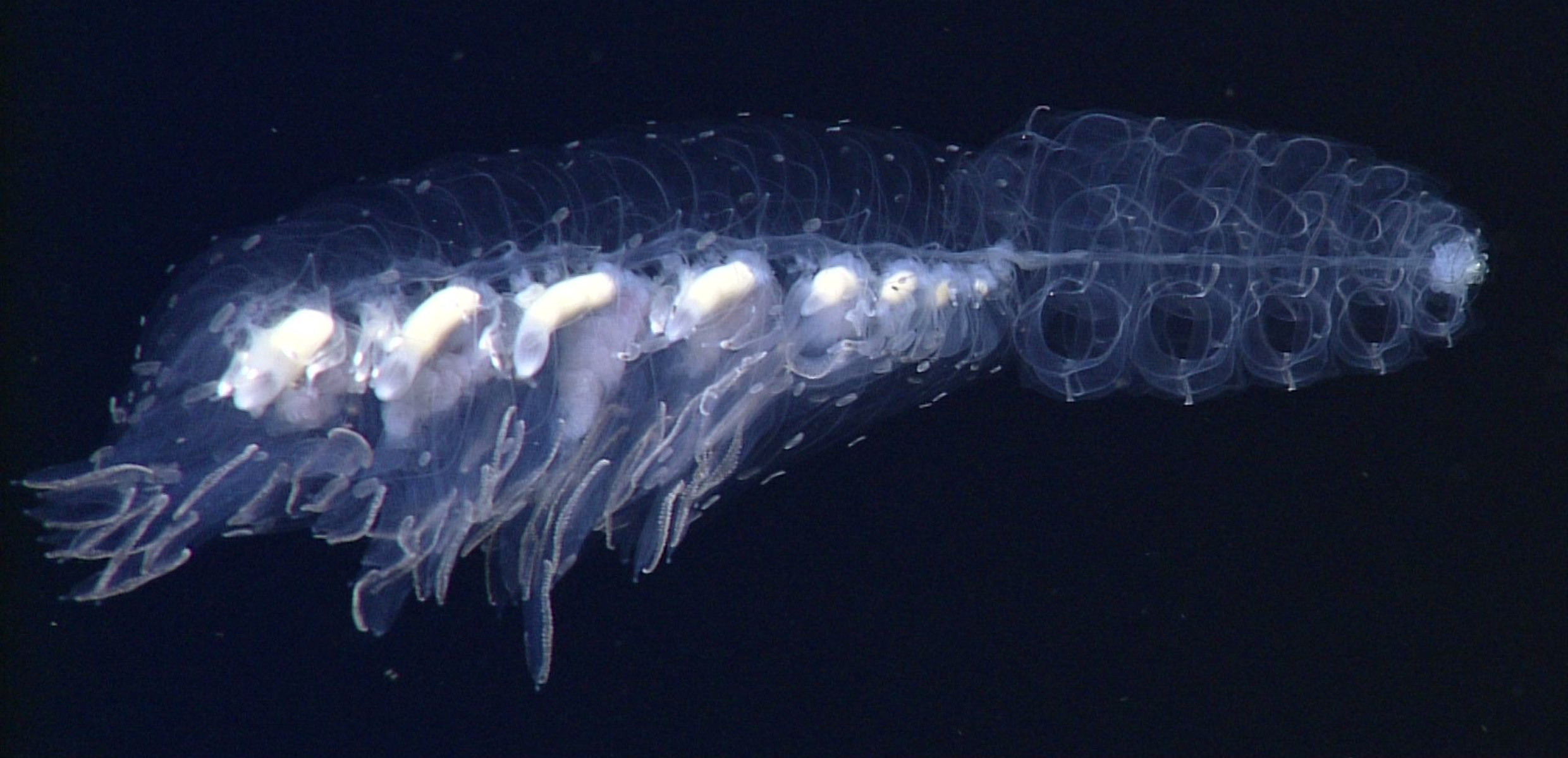
Description. An in situ frame grab of the type specimen is shown in Figure 26 View FIGURE 26 , and a more detailed one of the recently collected specimen from Doc Ricketts Dive 779 is shown in Figure 27 View FIGURE 27 .
Pneumatophore and Nectosome: In situ the nectosome of the type specimen occupied over half the total length with c. 29 mature nectophores and a number of nectophoral buds, together with the anterior pneumatophore. However, this region, having become denuded of its larger nectophores, does not appear to have been preserved.
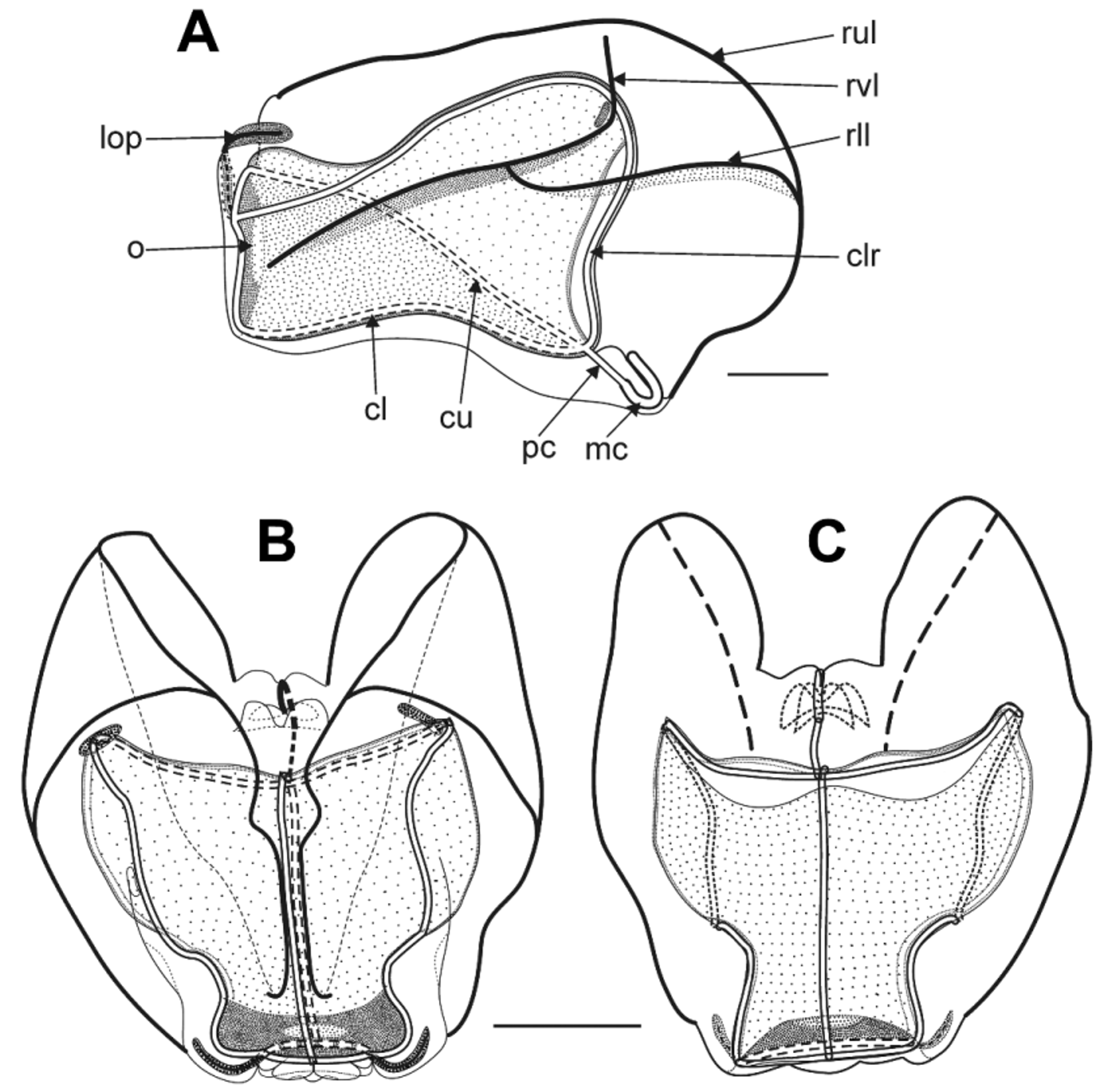
Nectophores: Twenty seven nectophores were found with the preserved type specimen that ranged in size from 12 to 30 mm in length and 6 to 29 mm in maximum width. The small nectophores ( Figure 28 View FIGURE 28 A) had yet to take on the flattened, in the upper/lower plane, appearance of the mature ones. The sides were folded up so that the upper lateral ridges ran parallel with each other and only a short distance apart. They defined the edges of a deep median furrow that was deepest proximally and gradually shallowed distally, as is indicated by the course of the upper radial canal in Figure 28 View FIGURE 28 A. Proximally the combined upper and lower lateral ridges originated in the region where the nectophore was attached to the stem. The latter branched from the former at about half the height of the nectophore, and each was divided into two sections, proximal and distal, with the distal part appearing as an extension of the vertical lateral ridge ( Figure 28 View FIGURE 28 A). At this stage, the vertical lateral ridge did not join with the upper lateral ridge and, at all stages, neither the upper nor lower lateral ridges reached the ostium, nor did the former divide. The vertical and lower lateral ridges stood out from the surface of the nectophore. A small, elongate patch of ectodermal cells was present on each side of the nectophore just distal to the vertical lateral ridge. In the preserved state it was brown in colour but, as with other Erenna species, almost certainly it was black in life. The nectosac was deeply divided in the mid-line by the upper furrow ( Figure 28 View FIGURE 28 A), but had extensive lateral wings in the proximal half of the nectophore. The mantle canal was thick and U-shaped, and gave rise to the short pedicular canal that, in turn gave rise to all four radial canals. For the most part the lateral canals followed the contours of the nectosac, except distally where they continued to the lateral margins of the ostium. All the canals were a brownishblack in colour in the preserved state, but almost certainly were black in life. There was a small muscle-free zone just distal to the lateral radial canals, where they ascended from their origin from the pedicular canal.
On the slightly larger nectophores ( Figure 28 View FIGURE 28 B, C) the upper lateral ridges still remained close together in the distal half of the nectophore, but the furrow between them was much shallower. The lower lateral ridge then appeared as a single entity and the vertical lateral ridges joined with the upper laterals. The muscle-free zone now consisted of a narrow band on either side of the proximal part of the lateral radial canals. The most interesting feature was the start of the development of the very characteristic thrust block. The central, proximal connection between the combined upper and lower lateral ridges, which formed the margins of the axial wings, consisted of two hemispherical mesogloeal thickenings that lay each side of a deep groove through which passed the ascending mantle canal. Distal to these, on the upper side of the nectophore, there were two further pairs of roughly hemispherical mesogloeal thickenings. The more distal pair was the larger and partially overhung the other pair.
On the fully developed nectophores ( Figure 29 View FIGURE 29 A, B) the upper lateral ridges had now moved out toward the lateral margins. As before they did not quite reach to the ostium. The lower lateral ridges branched from the upper ones on the lateral margins of the axial wings, and these too did not quite reach ostial level. The vertical lateral ridges were distinct and there were small patches of, presumably, cells producing bioluminescence on the distal margins. Proximally the apex of the nectosac was almost flat and there was a clear muscle free zone, mainly on the lower side, but occasionally extending on to the upper one. The thrust block had expanded considerably and now formed two rounded lobes on the upper side of the nectophore. Below these and extending further proximally, was the pair of rounded thickenings that were deeply divided in the mid-line with the ascending mantle canal running through the furrow. The lateral ostial processes were well developed and even by the mature stage still contained nematocysts that appeared to be isorhizas ( Figure 29 View FIGURE 29 C).
Siphosome: Because of the highly contracted nature of the preserved stem and the dense clusters of zooids, it was not possible to glean any useful information concerning the organisation of the individual cormidia.
Bracts: Although very similar, the bracts could be divided into two types. The Type A ( Figure 30 View FIGURE 30 A) were slightly more abundant than the Type B ones ( Figure 30 View FIGURE 30 B). They were elongate, measuring up to c. 45 mm in length and 20 mm in width. They bore a pair of asymmetrically placed lateral cusps in their proximal half, with the cusp on the inner side usually being the more distal of the pair and more pronounced. There was a small circular patch of ectodermal cells on the upper side in the mid-line slightly distal of the mid-length. The inner side, at the proximal end, was distinctly thickened with mesogloea so that the bracteal canal ran over a sort of keel. The canal arose on the upper side of the bract and shortly after passing on to the lower side it usually thickened slightly. It continued distally and in contact with the lower wall until close to the distal end. There it thinned as it penetrated into the mesogloea and ran diagonally upwards to reach the upper surface toward the proximal end of a distinct raised patch of ectodermal cells, and then continued distally for a short distance immediately below the patch. Throughout its length the canal contained, in the preserved state, dark brown pigmentation that probably was black in life.
This distal patch of cells ( Figure 31 View FIGURE 31 ) was roughly rectangular in shape with rounded corners. Its upper side consisted of a honeycomb of ectodermal cells interspersed with a few nematocysts. This probably was the result of abrasion as the more protected sides of the patch were packed with nematocysts. These nematocysts ( Figure 31 View FIGURE 31 , insert) measured c. 100 x 28 µm and were probably of the same type that were found on the lateral ostial processes of the nectophores. However, no evaginated ones were observed.
The bracts of the second type ( Figure 30 View FIGURE 30 B) were shorter, measuring up to 25 mm in length and 17 mm in width. They also possessed a pair of lateral cusps in their proximal half, but in this case the cusps were almost symmetrically arranged. Again, there was a small central patch of ectodermal cells, and the course of the bracteal canal was the same as for the other type of bract. However, on the inner side of the bract, at the proximal end, the mesogloea was uniformly thickened and so there was no pronounced keel. In the very youngest bracts there was a conical protuberance on one side of the bract, proximally, but this disappeared as the bracts enlarged. The distal ectodermal thickening on the upper side of the bract was basically the same as for the other type of bract, although in general it was less extensive.
Gastrozooid and tentacle: The large gastrozooids ( Figure 32 View FIGURE 32 ) were largely black in colour and, in both life and after preservation, had an asymmetrical shape. The small, ring-like basigaster was attached directly to the stem and was separated from the main stomach region by an annulation. The stomach was usually laterally expanded and the distal proboscis region arose at an oblique angle on one side. The distal region was paler than the others and numerous endodermal, longitudinal "hepatic stripes" could be seen. The tentacle arose from one side of the basigaster, and was annulated and bore strong musculature.
Tentillum: The tentillum ( Figure 33 View FIGURE 33 ) was of a unique morphology, although typical for an Erenna species. It consisted of an obviously highly extensile, but in its preserved state a highly contracted pedicle, as exemplified by the corrugated nature of the gastrovascular cavity. The remainder of the tentillum was a cylindrical tube, but flattened on its inner side. It was U-shaped in its preserved state and measured c. 55 mm in length, but in life was quite straight (see Figure 27 View FIGURE 27 ). Its ectoderm consisted of irregularly shaped small cells, below which was an amorphous layer of mesogloea, and then a spongy internal layer of pentagonal or hexagonal-shaped cells surrounding the narrow gastrovascular cavity ( Figure 33 View FIGURE 33 B). This spongy layer decreased in size distal to the cnidoband itself, forming a narrower, short terminal process that terminated in a spherical photophore ( Figure 34 View FIGURE 34 D). There the gastrovascular cavity gave rise to a diverticular canal that ran proximally, above the main gastrovascular cavity to the middle of the cnidoband ( Figure 33 View FIGURE 33 C, D). Whether it then gave rise to two branches that ran distally and proximally along the base of the cnidoband could not be discerned. Young tentilla, at various stages in development are shown in Figure 34 View FIGURE 34 A. The outer process that forms the cnidoband is distinct at a very early stage in development. No details with regard to the ocellus and its potential luminescent properties are presently available.
The cnidoband itself, as with other Erenna species (see Pugh, 2001) was a discrete structure that measured c. 12.5 mm in length in its preserved state ( Figure 33 View FIGURE 33 B, C). The nematocysts were concentrated around the outer lateral margins of the cnidoband in a much folded sequence giving the outer edge a scalloped appearance. The large nematocysts, presumably microbasic mastigophores ( Figure 36 View FIGURE 36 B), which measured c. 142 x 23 µm, were present in relatively large numbers compared with the other Erenna species. The smaller ones ( Figure 34 View FIGURE 34 C), which were quite variable in size, measuring c. 39– 42 x 16–20 µm, appeared to be all of the ovoid, pear-shaped type. No discharged ones of either type were found. Whether the small cylindrical type of nematocyst was absent or so rare that it was not noted was unclear.
Palpon: Each palpon ( Figure 35 View FIGURE 35 ) consisted of a short pedicle, a slightly broader part, separated from the more distal parts by a constriction, to which the palpacle was attached. The latter was often lost, but when present, was very short and possessed no nematocysts. The main part of the palpon consisted of a large, flimsy bag-like structure, together with a distinct, narrow, whitish proboscis region. The main body was covered in an array of long irregular patches of large ectodermal cells interspersed with or overlaying more homogeneous smaller cells. The largest palpons measured up to c. 20 mm in length and 10 mm in diameter. Groups of palpons arose from a common pedicle and these were usually closely associated with the gonodendra. However, the highly contracted nature of the preserved stem did not allow for the exact arrangement to be elucidated.
From 30 to 50 nematocysts were found around the distal mouth opening that measured c. 92 x 28 µm ( Figure 35 View FIGURE 35 inset). Although of similar size to those found on the nectophores and bracts they did not appear to be of the same type. Possibly they were mastigophores, but this could not be confirmed as no discharged ones were found.
Gonodendron: The type specimen was male and gonophores, in various stages of development, were found attached to several gonodendra ( Figure 36 View FIGURE 36 A). There appeared to be more than one of these per cormidium, but due to the highly contracted state of the preserved specimen, this could not be verified with certainty. Each gonodendron appeared to be associated with a cluster of palpons, but again the highly contracted nature of the preserved siphosome did not allow an accurate assessment of their disposition. The younger gonophores were clustered closely together, but as they matured they became more separated, being borne on rigid, brittle, almost twig-like, stalks.
Each gonophore, up to c. 3.5 mm in diameter, was a spherical, bag-like structure with a somewhat amorphous but spherical, central manubrium containing the sexual products that occupied only part of the subumbrella space. Typically four straight radial canals could be discerned, but they were not arranged symmetrically (see Figure 36 View FIGURE 36 B). Their arrangement suggested that there should have been five canals, spaced approximately 72° apart, but with one canal missing. Occasionally (see Figure 36 View FIGURE 36 B) this fifth canal was present, while on rare occasions there were only three canals that reached the ostial ring canal. The ostium was completely closed by a circular membrane comprised of an outer ring of large, roughly circular cells surrounding smaller cells.
Distribution. Known from only two specimens, one collected from a depth of 3098 m at 34°17.23'N, 124°03.10'W, west of Point Conception, California, where the water depth was c. 3450 m, and the other west of the Davidson Seamount, California at a depth of 2800 m, where the water depth was c. 3770 m.
Etymology. The specific name insidiator is derived from the Latin insidia, and means an ambusher as, although not proven, its tentillum probably, in some way, acts as a lure.









No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.