
Eurytoma erythrinae Gates and Delvare, 2008
|
publication ID |
https://doi.org/ 10.11646/zootaxa.1751.1.1 |
|
persistent identifier |
https://treatment.plazi.org/id/03B1913B-482F-2034-FF3A-5A7CFDADFF69 |
|
treatment provided by |
Felipe |
|
scientific name |
Eurytoma erythrinae Gates and Delvare |
| status |
sp. nov. |
Eurytoma erythrinae Gates and Delvare , new species
(Figs. 1–16)
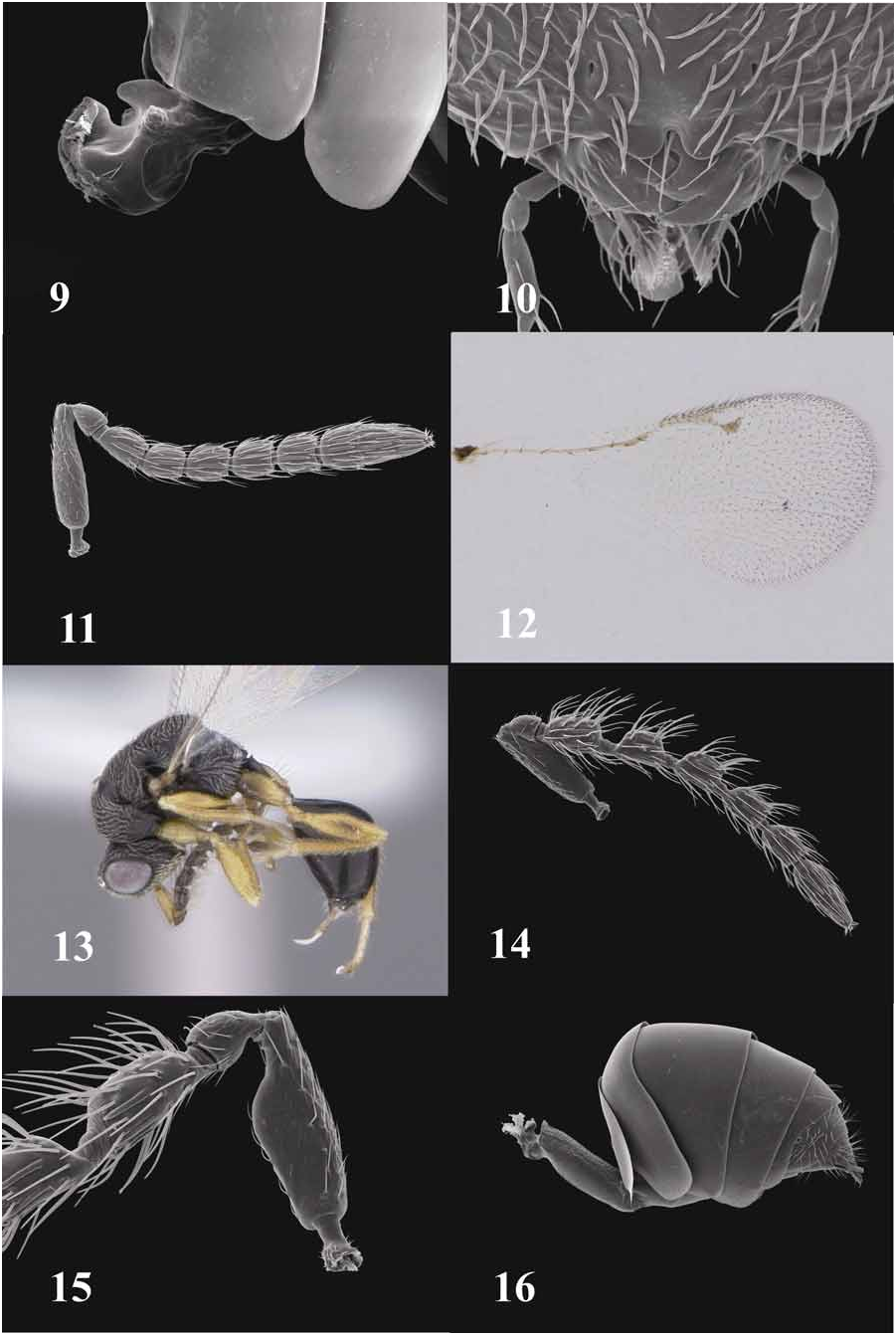
Female holotype. Body length 2.4 mm. Color: Black except for the following straw yellow—scape, pedicel mesally and apically, anterolateral spot on pronotum, pro- and mesocoxae, front leg entirely, meso- and metatibiae, meso- and metafemora, and ovipositor sheaths (Fig. 1); fuscous—pedicel laterally and dorsally, flagellum, tegula, profemoral spot laterally, metafemoral band mediolaterally, and metacoxa; whitish—tarsomeres and wing veins.
Head 1.4X as broad as high, punctured with short radiating striae originating from oral fossa; anterior tentorial pits small but present; genal carina present, gena effaced-microreticulate, outline in frontal view straight on upper half and slightly convex below (Fig. 2); malar space 0.71X eye height, asetose; scrobal basin cari- nate laterally, carina raised into a short lobe dorsad toruli and fading apically (Fig. 2); mandible tridentate, basal tooth truncate and separated from middle tooth by broad emargination; clypeus bilobate and supraclypeal area smooth ( Fig. 10 View FIGURES 9–16 ); toruli positioned slightly above lower ocular line, inner edges distinctly raised; intertorular space sulcate and bearing two rows of hairs, not pointed above. Scrobal depression carinate laterally, carina raised as a lobe dorsad toruli and fading apically (Fig. 2). Antenna with scape reaching ventral margin of anterior ocellus; ratio of scape (minus radicle):pedicel:anellus: F1:F2: F3:F4:F5:club as 33:8:1:8:7:7:7:6:19; pedicel chalice-shaped; funicle fusiform; F1 slightly narrowed basally; funicular segments with single row of longitudinal sensilla ( Fig. 11 View FIGURES 9–16 ) and two whorls of setae, the latter slightly shorter to as long as its bearing segment; clava bisegmented. Ratio of lateral ocellus:ocellocular distance:postocellar distance as 6:5:21. Head posteriorly lacking postgenal lamina but postgenal grooves evidently ridged, not converging in their lower part, extending ventrally to upper margin of hypostomal bridge; dorsal margin of lateral foraminal plate visible, convex; subforaminal plate absent; postgenal sulci narrow, superficial. Mesosoma umbilicate (Fig. 3), 1.80X as long as broad; midlobe of mesoscutum 1.17X as long as broad; scutellum 1.33X as long as broad; notauli complete, shallow; axillar grooves with pit at mid length; lateral surface of prepectus triangular, broadly rounded posteriorly, smooth; subventral carinae of prepectus Y-shaped, joined posteriorly, ventral surface of prepectus without median tooth. Mesepimeron reticulate ventrally, dorsally delimited from femoral depression by fine carina with femoral depression reticulate at mid-height. Mesepisternum anterior to femoral depression umbilicate (Fig. 7), epicnemium imbricate, flattened, with superficial submedial, shallow depressions to receive procoxae (Fig. 7) [discrimen visible as anteromedial ovate depression]. Metapleuron and lateral areas of propodeum (Figs. 4, 5) shallowly umbilicate, propodeum broadly flattened, medially smooth, bordered mediolaterally by numerous carinae forming irregular asetose cells, these bordered laterally by setose cells (Fig. 5), basal carina clearly visible, raised into small teeth at anterior corners of median groove; cluster of setae anterolaterad nucha; spiracle about 1/3 its greatest diameter from dorsellum. Procoxa imbricate except anterior surface smooth, superficially depressed anteriorly for reception of lower head, the depression margined dorsally by very faint ridge (Fig. 6). Mesocoxa lacking lamella. Metacoxa elongatereticulate anteriorly, becoming reticulate posteriorly, lacking dorsal hairs basally. Forewing with ratio of marginal vein:postmarginal vein:stigmal vein as 21:15:12 ( Fig. 12 View FIGURES 9–16 ). Metasoma smooth, Gt 6 and syntergum weakly microreticulate (Fig. 8); petiole 0.7X as long as broad in dorsal view, with projecting lateral teeth as well as mediodorsal prong ( Fig. 9 View FIGURES 9–16 ); transverse ridge between petiole and Gs1 absent; gaster teardrop-shaped in lateral view, ovipositor angled at about 30° dorsad horizontal axis (Fig. 8); measurements of gastral terga along midline as 15:16:26:35:8:5:12; Gt4 emarginate posteriorly in dorsal view; Gt6 parabolic in cross section.
Male. Body length 1.9 to 2.3 mm (n=7). Color: black, yellow areas as described for female ( Fig. 13 View FIGURES 9–16 ). Sculpture as described for female. Antennal with funicular segments ( Fig. 14 View FIGURES 9–16 ) pedicellate, each with 2 or more rows of erect setae and about 1.5X as long as width of segment; ratio of scape (minus radicle):pedicel:anellus:F1:F2:F3: F4:F5:club as 21:7:1:12:11:11:11:10:18; scape with ventral plaque in apical half ( Fig. 15 View FIGURES 9–16 ), cellular structures visible beneath cuticle. Gastral petiole in lateral view cylindrical ( Fig. 16 View FIGURES 9–16 ), in dorsal view length about 3 X as long as greatest width, subequal in length to metacoxa; evenly reticulate dorsally and ventrally, smooth laterally.
Variation. Females vary in length from about 2.7 to 3.0 mm. In females, the fuscous area on the profemur can form a band in the middle half of the femur (but discontinuous medially), the gena adjacent to the malar carina can tend toward being completely smooth with microreticulation almost completely effaced. The epicnemial carina is sometimes incomplete in the anterior half. The procoxa may be marked with dark brown on the anterior surface. The pro- and mesocoxa may be entirely dark brown. The South African specimens (Nelspruit Bot. Garden) have shallower setigerous punctation overall and a more finely punctate, less defined median channel on the propodeum. Some South African males have a more yellowish coloration to the facial setation, and the hind femur can range from golden to having a brownish band in the medial half. A male from Ghana displays a condition similar to material from the Nelspruit Bot. Garden in terms of sculpture reduction, and its metafemur is almost entirely brown.
Type specimens. Holotype, ♀, TANZANIA: Arusha: Maroron Village , 24.i.2006. M. Ramadan, emerging from Erythrina abyssinica galled seedpods, T2 -B (USNM). Paratypes (26♀ 17♂): 8♀ 9♂, same data as for holotype (2♀ 3♂, USNM; 2♀ 1♂ BMNH) and same data as for holotype except T2 - C (2♀ 3♂, ANIC; 2♀ 2♂ CIRAD) . SOUTH AFRICA: Durban Harbor, R40 , 8.IV.2006, coll. M. Wright, R . Messing, D. Rubinoff; emerged from galled leaves of Erythrina lysistemon , EGW-17 (2♀ 1♂ BPBM); Along Road R40 , 1.IV.2006, coll. M. Wright, R . Messing, D. Rubinoff; emerged from galled leaves of Erythrina lysistemon , EGW-4 (6♀ 1♂ SAMC); Nelspruit, Botanic Garden , 1.IV.2006, coll. M. Wright, R . Messing, D. Rubinoff; emerged from Erythrina lysistemon galls, EGW-10 (4♀ SAMC) . GHANA: Adomi Mamiwata, 7. V .2006, coll. A. Bokonon- Ganta, emerged from galls on Erythrina sp. , 6 B2-A/41 (1♀ 1♂ FSCA); Aneho , 30.IV.2006, coll. A. Bokonon- Ganta; emerged from galls on Erythrina vogelii , 12 B2-L/66 (1♀ NCSU) .
Etymology This epithet is a noun in apposition, feminine singular, and named for Erythrina (Fabaceae) , the genus of plant with which it is associated.
Host. Reared from field populations of the gall-forming Quadrastichus spp. Note: There appear to be more than one species of Quadrastichus associated with galls from which E. erythrinae emerged (J. LaSalle, pers. comm.).
Distribution. Ghana, Tanzania, and South Africa.
Discussion. The bilobed clypeus, habitus of the intertorular space, fusiform funicular segments, Y-shaped subventral carinae of the prepectus, flat epicnemium, ornamentation of the propodeum, long gastral tergite 4 indicates that E. erythrinae is a member of the dentata species group as defined by Lotfalizadeh et al. (2007). However, it differs from most species in this group by lacking produced postgenal laminae that converge ventrally and a procoxal carina anteriorly, though there is a depression. Most members of the dentata- species group, where biology is known, attack Cecidomyiidae ( Lotfalizadeh et al. 2007) . Eurytoma dentata Mayr (1878) was reared from several gall-midges ( Bouček 1977), especially of the genus Asphondylia ( Parnell 1964, Noyes 2003). Another species of the same group was reared in Senegal from another Asphondylia galling ovaries of the plant Solanum aethiopicum L./Torner ( Solanaceae ) ( Etienne & Delvare 1987). However, at least one species of the group is phytophagous, the larvae developing in pods of Haematoxylon campechianum L. ( Fabaceae ) in Guadeloupe (J. Étienne, pers. comm.). Examination of type material of Eurytoma species described from Africa by Risbec (1951, 1952, 1953, 1955, 1956, 1957a, 1957b) showed that it most closely resembles E. radicicola Risbec (1952) , but is differentiated as follows (condition in E. erythrinae in parentheses): E. radicicola with moderately converging genae, straight in frontal view (convex in frontal view), elevated genal carina (elevation not discernible), F2–F5 longer than broad (subquadrate), median channel of propodeum delimited by carina laterally (poorly delimited), procoxa with oblique carina anteriorly (carina very faint), marginal vein 2X as long as postmarginal vein (subequal in length), and gastral profile straight along terga 3 and 4 (regularly convex). Eurytoma radicicola was collected in Madagascar from galls on the roots of orchids ( Risbec 1952).
Classification. Based upon the recent morphological phylogenetic analysis of Lotfalizadeh et al. (2007), several genera have been re-characterized. Not surprisingly, the speciose genus Eurytoma proved polyphyletic and the genus was redefined based upon four putative synapomorphies. Eleven species groups are included in their Eurytoma s.s. Other genera were redefined based upon synapomorphic support and numerous nomenclatural changes proposed in Lotfalizadeh et al. (2007). Those changes proposed in the checklist below are based upon the redefined generic concepts of Lotfalizadeh et al. (2007), but were beyond the scope of that particular research effort.
Lotfalizadeh et al. (2007) included the dentata species group in the Phylloxeroxenus clade, which is species-rich and highly diverse in tropical regions. Unfortunately, their data do not support a robust generic classification due to instability in the trees based upon their morphological phylogenetic analysis. Further studies are needed, especially using molecular data, for a more robust phylogeny and to provide support for a new generic classification. Pending such a study, no alternate genera are available to receive E. erythrinae ; thus, our placement in Eurytoma , even though it does not fit the narrower concept of the dentata species group as proposed by Lotfalizadeh et al. (2007).
| USNM |
Smithsonian Institution, National Museum of Natural History |
| ANIC |
Australian National Insect Collection |
| CIRAD |
Centre de Cooperation Internationale en Recherche Agronomique pour le Developpement |
| R |
Departamento de Geologia, Universidad de Chile |
| BPBM |
Bishop Museum |
| SAMC |
Iziko Museums of Cape Town |
| V |
Royal British Columbia Museum - Herbarium |
| FSCA |
Florida State Collection of Arthropods, The Museum of Entomology |
| NCSU |
North Carolina State University Insect Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |