
Liolaemus cyaneinotatus, Martinez, Lorena Elizabeth, Avila, Luciano Javier, Perez, Cristian Hernan Fulvio, Perez, Daniel Roberto, Sites, Jack W. & Morando, Mariana, 2011
|
publication ID |
https://doi.org/ 10.5281/zenodo.204374 |
|
DOI |
https://doi.org/10.5281/zenodo.5622934 |
|
persistent identifier |
https://treatment.plazi.org/id/2262D172-FFD1-9E23-FF49-FDB7FBA7F0A6 |
|
treatment provided by |
Plazi |
|
scientific name |
Liolaemus cyaneinotatus |
| status |
sp. nov. |
Liolaemus cyaneinotatus sp. nov.
Type material. Holotype. — MLP.S 2618 ( Fig. 1 View FIGURE 1 ), an adult male from Auca Mahuida (37° 41’ S, 68° 48’ W, 1332 m), Pehuenches Department, Neuquén Province, Argentina, C. Perez collector, 28 March 2008.
Paratypes.— MLP.S 2619 to 2622, LJAMM-CNP 10381, 10388 (females), LJAMM-CNP 10382-85, 10387 (male) (37° 41’ S, 68° 48’ W, 1332 m), C. Perez collector, 28 March 2008; LJAMM-CNP 10389 (male) (37° 43’ S, 68° 53’ W, 1757 m), C. Perez collector, 28 March 2008; LJAMM-CNP 10551 (male) and LJAMM-CNP 10552 (female), from Auca Mahuida (37° 43’ S, 68º 55’ W, 1983 m), C. Perez collector, 28 March 2008. All localities are on Auca Mahuida Volcano, Pehuenches Department, Neuquén Province, Argentina.
Diagnosis. Liolaemus cyaneinotatus is a small, slender lizard, included in the alticolor - bibronii group ( Lobo et al. 2010). The new species differs from L. araucaniensis , L. bibronii , L. curicencis , L. fuscus , and L. lemniscatus in having conspicuous cyan spots irregularly distributed in dorsal and lateral areas of the body. Liolaemus araucaniensis has a greenish background coloration, with well developed but irregular and sometimes fused dorsolateral transversal marks, some fused with the vertebral line, a pattern never present in L. cyaneinotatus . Liolaemus cyaneinotatus is a slightly smaller species than L. curicensis (maximum SVL 59.0 mm vs 64.2 mm) and L. araucaniensis (maximum SVL 60.9); both species have higher mean of snout-vent length (60.9 mm and 60.4 mm vs. 51.0 mm) and higher mean of distance from axilla–groin (27.2 mm and 28.8 mm vs. 21.7 mm). Liolaemus araucaniensis , L. curicensis and L. lemniscatus have higher mean tail length than L. cyaneinotatus (104.3 mm, 112.5 mm, and 105.0 mm vs. 82.3 mm). Vertebral line is absent in Liolaemus lemniscatus unlike the new species. Liolaemus cyaneinotatus differs from L. araucaniensis , L. curicensis , L. fuscus , L. gracilis and L. lemniscatus in having a higher maximum number of scales around midbody (68 vs. 58, 56, 53, 44 and 58 respectively); L. curicensis has a greater maximum number of fourth toe infradigital lamellae on the hind foot (27 vs 24) ( Table 1 View TABLE 1 ). Liolaemus cyaneinotatus differs from L. gracilis in having higher mean number of scales in the following characters: dorsal scales (62.2 vs. 51.2), dorsal head scales (12 vs. 11.5), scales around interparietal (6.6 vs. 5.9), enlarged supraoculars (6.1 vs. 5.9), number of temporals (7.2 vs. 6.4), number of gulars (28.9 vs. 25.3), scales around nasal (6.8 vs. 6.4) and infradigital lamellae of 4th toe of the hind foot (23.1 vs. 21.8). Liolaemus gracilis have always well marked dorsolateral white stripes and usually lacks of paravertebral markings. Liolaemus bibronii have always well marked black vertebral and dorsolateral lines, as well as a pattern of paravertebral markings well defined and larger than L. cyaneinotatus ; never have brilliant cyan scales dispersed in dorsal and lateral areas, and general background coloration is always dark, almost black in some individuals, without orange or yellow-orange clear coloration as L. cyaneinotatus . Mean of scales counts are higher in L. cyaneinotatus than L. bibronii in: dorsal head scale (10.75 vs. 13.07), enlarged supraoculars (3.95 vs. 4.60), scales between rostral and frontal (4.84 vs. 5.40) and infradigital lamellae of 4th toe of the foot (21.93 vs. 23.07). Liolaemus cyaneinotatus have lower mean number of gulars scales (30.29 vs. 28.67), neck scales (16.27 vs. 14.67), and loreolabials scales (6.40 vs 5.60) than L. bibronii (Tables 2 and 3).
Description of holotype. Adult male ( Fig. 2 View FIGURE 2 ): snout–vent length (SVL): 55.4 mm, tail length (TL): 78.29 mm regenerated, axilla–groin distance (DFH): 25.01 mm, head length (HL): 12.29 mm, head wide (HW): 9.69 mm, humerus length (HUL): 6.29 mm, radius-ulna length (RUL): 5.46 mm, hand length (HAL): 7.76 mm, femur length (FL): 8.12 mm, tibio-fibula length (TFL): 10.70 mm, foot length (FOL): 15.19 mm, fourth-toe length (4TL): 13.04 mm, maximum body height (HMAX): 6.99 mm, maximum body width (WMAX): 13.83 mm.
Character cyaneinotatus n. sp. araucaniensis curicencis continued.
Character fuscus gracilis lemniscatus
n=40 n=47 n=97
Snout–vent length 49.6 (48.2–50.5) 48.0 (39.4–57.3) 51.3 (45.8–55.7) Distance from axilla–groin 22.4 (21.8–23.6) 22.1 (17.3–28.1) 23.7 (21.0–27.0) Vertebral line present present absent Tail length 85.6 (79.0–90.0) 92.8 (72.2–110.2) 105 Scales around midbody 47–53 34–44 50–58 Infradigital lamellae of 4th toe 22–25 20–26 23– 24 Male precloacal pores 2–3 3–4 2– 3 Females with precloacals pores absent absent absent Reproductive mode oviparous oviparous oviparous Elevational range 1600 0–1400 1800
Dorsal head scales smooth, 10 from a line drawn horizontally between anterior margin of external auditory meatus to anterior margin of rostral. Left nasal scale in contact with rostral. Two postrostral scales and four internasals. Canthals separated from nasal by one scale. Five enlarged supralabial scales, the fourth (2.54 mm) with the posterior margin curved upward but not contacting subocular. Four infralabial scales. Auditory meatus oval (height: 2.39 mm; width: 1.31 mm). Scales of anterior margin of auditory meatus without keels, scales of the posterior margin smaller and granular. Three enlarged, flat scales projecting over meatus; no auricular scale differentiated. Seven convex, imbricate and slightly keeled temporal scales between upper anterior corner of auditory meatus and posterior margin of orbit. Six temporals (counting vertically from bucal commisure to level of supercilliaries). Interparietal scale subpentagonal, bordered by six scales. Parietals and interparietal of similar size. Orbitauditory meatus distance (4.46 mm), almost twice to orbit-rostral distance (anterior margin of rostral: 7.01 mm). Rostral scale about twice wider than high (width: 2.66 mm; height: 1.23 mm). Mental scale subpentagonal, also almost twice wider than high (width: 2.58 mm; length: 1.50 mm) in contact with first infralabial (on each side). Frontal scale not divided. Five scales between frontal and rostral, five scales between frontal and supercilliaries. Four enlarged supraoculars. Five strongly imbricate supercilliaries scales. Subocular longer (3.93 mm) than orbit diameter (2.34 mm from anterior to posterior margin of cilliaries) and separated from supralabials by a single row of lorilabials. Six lorilabials, with fourth through sixth contacting subocular. Preocular in contact to lorilabials. Postocular elongated, level with fifth supralabial scale. Posmental row formed by three enlarged scales, second posmentals separated from each other. Scales of throat flat and imbricate. Twenty-seven gulars between auditory meatus. Lateral side of the neck flat, formed by small scales, imbricate and keeled. Dorsal scales lanceolate, imbricate and moderately keeled. Forty-nine scales around midbody. Sixty-three dorsal scales between occiput and anterior level of thigh. Ventral scales larger than dorsals (dorsal scale length: 1.22 mm; ventral scale length: 1.39 mm). Eigthy ventral scales counted at the midline between rostral scale and vent. Three precloacal pores. Seventeen subdigital lamellae in the fourth toe of the hand and twenty-four on the foot. Anterior suprabrachials rhomboidal, imbricate, smooth, slightly larger in size to dorsal body scales. Postabrachials smaller, smooth, becoming granular near axilla. Supra-antebrachials similar to suprabrachial. Pre-antebrachials imbricated, rhomboidal, keeled. Infraantebrachials rhombals, imbricate, keeled. Supracarpals imbricated, rhomboidal, smooth. Infracarpals strongly imbricate, rhomboidal, keeled, mucronate. Subdigital lamellae with 3 keels, each terminating in a short mucron, numbering: I: 9, II: 12, III: 15, IV: 17, V: 12. Claws robust, curved and sharp, opaque brown.
Suprafemorals as large as dorsal body scales, rhomboidal, imbricated, slightly keeled near the insertion to strongly keeled and terminating in a short mucron. Infrafemorals and prefemorals rhomboidal, imbricated, smooth. Postfemorals small, granular shape. Supratibial rhomboidal, imbricated, strongly keeled and with a short mucron. Infratibials rhomboidal, imbricated, smooth. Supratarsals rhomboidal, imbricated, smooth with some scales slightly keeled in the outer side. Infratarsal small, rhomboidal, imbricate, keeled, mucronate. Subdigital lamellaes of fingers with three blunt keels each terminating in 3 short mucrons, numbering: I: 10, II: 14, III: 22, IV: 23, V: broken. Claws robust, curved and sharp, opaque brown. Tail quadrangular in cross section near cloacal area, becoming oval to round towards the tip. Dorsal and upper lateral caudal scales keeled, imbricate, mucronate. Lower lateral scales and ventral scales weakly keeled.
Color in life. Head predominantly light brown, darker than body; some scales with a brick-orange tinge. A distinctive white line between postocular scale and antehumeral fold, a scale wide, black outlined. Subocular scale white, with black upper border. Dorsal background coloration is ferruginous orange. Dorsal pattern with a tenuous light brown vertebral field 2–3 scales wide at midbody. Vertebral line black, well marked, continuous from occiput to first third of the tail, becoming broken to the tip. Paravertebral field, four-five scales wide, slightly darker than vertebral field, with dark ferruginous orange. Paravertebral markings black, irregulars, a scale size; eight series between shoulders and pelvic girdle, not paired. Between occiput and shoulder, paravertebral markings fused, forming two continuous black lines; after pelvic girdle becomes regularly paired and fused to the tip of the tail. Dorsolateral stripes whitish, one scale wide, originating on the posterior border of each orbit, and disappearing in the first third of trunk. Iridescent cyan scales scattered at midbody following the dorsolateral stripes and contact area between paravertebral and lateral fields. Lateral field with a light orange background slightly darker than paravertebral field, speckled with black, white, and iridescent cyan scales ( Fig. 1 View FIGURE 1 ). Lateral field coloration progressively fades ventrally, where light gray replaces the orange background coloration, except in the proximity of the hind limbs insertions were the orange coloration becomes more intense. Throat background light gray, with a black reticulated pattern. Chest, belly, tail, and ventral surfaces of the thighs light gray. Lower belly, cloacal apron, ventral femoral area, and first third of the tail, yellow-orange. First lateral portion of the tail bright orange. Precloacal pores bright orange.
Color in preservative. Coloration pattern is maintained but background coloration becomes darker. All cyan dots disappear as well as all orange or yellow coloration. Ventral coloration becomes gray, more intense in the middle of each scale.
Variation. Table 4 View TABLE 4 summarizes morphometric and scale variation data. In this section we provide information on traits not included in the table. Dorsal head scale surface nearly uniform, smooth (or slightly wrinkled in seven specimens); temporal scales keeled in five, smooth in ten. Fourth supralabial scale is curved upward at its posterior margin in all except five specimens, and only contacts the subocular in only five individuals. Three individuals with nasal scale separated from rostral. Posterior half of supraorbital semicircles complete in thirteen, incomplete in two. Canthal scale separated from nasal by two scales, except one specimen separated by one scale. Frontal scale divided in three, undivided in twelve. Vertebral line varies from well marked to completely absent. Paravertebral field disappear completely in some individuals. Paravertebral marks become fused in some individuals forming a line equal to the vertebral line, or are reduced to small dots or disappear in some individuals. Some males have a dorsal plain pattern alive, without vertebral or paravertebral lines or paravertebral marks. Dorsal background coloration varies from light gray to brick orange. Lateral field varies in melanism according to the density of black scales. Cyan dots are present in adult males, but can disappear in subadults. Density and distribution of cyan dots varies between individuals but always are in the lateral and/or dorsolateral areas of the trunk. Usually are absent in females. Yellow coloration is usually present in ventral areas of femurs, cloacal apron, first third of the tail and lower belly and varies in intensity from individual to individual becoming more extensive in females that in males. In males usually predominates yellow that orange coloration. Brick orange coloration is more intense in females and is stronger in lateral sides of the trunk, lateral areas of the first third of the tail, and prefemoral areas. In some females orange brick coloration form a continuous line between postoculars and antehumeral fold, upper part of the lateral field between axilla and prefemoral region, and from postfemoral area to the tip of the tail.
Etymology. Specific epithet is in reference to the cyan dots present in adult males in dorsolateral areas of the body, (cyanei = cyan; notatus = dotted).
Geographic distribution. Liolaemus cyaneinotatus is known only from the type locality where the holotype and paratypes were collected in rocky patches or open substrate outcrops above 1500 m elevation on Auca Mahuida volcano, Añelo Department, Neuquén Province, ( Fig. 3 View FIGURE 3 ). Auca Mahuida Volcanic Field is part of the Payenia, a large Quaternary volcanic province of basaltic composition in the foreland region, behind the active volcanic arc, unique in the entire Andean Chain ( Ramos & Folguera 2011). The Auca Mahuida Volcanic Field is an isolated basaltic plateau with the Auca Mahuida volcano in its center, surrounded by dunes fields or sandy flatlands with extremely arid characteristics, and is an island of Patagonian-like environment in the middle of Monte phytogeographic region. Liolaemus cyaneinotatus is not in contact with the geographic distribution range of other members of the L. bibronii group, with the exception of L. gracilis , found in lower altitudes of the Auca Mahuida volcano. But no sympatry was observed between these two species.
Natural history. The individuals were found in active basking in the edges of shrubs ( Prosopis denudans, Senna kurtzii, S. arnottiana) or on the top of rocks. Individuals were found active between 09:00 to 19:00 in sunny days of austral summer. Activity starts with basking behavior along the edges of the bushes or rocks. Several individuals were observed sharing basking areas and usually juveniles and adults are found under small bushes, sharing refugee or foraging areas. They usually foraged actively in the bushes but use the sand edges or tip of branches to bask. They seem to be territorial at some point because aggressive encounters between large males or females were observed. This species shares its habitat with other Liolaemus species ( Liolaemus austromendocinus , Liolaemus aff. elongatus , Liolaemus aff. boulengeri , Phymaturus sitesi and P. roigorum ). Liolaemus cyaneinotatus usually occupies the microhabitat around the shrubs that grow scattered separated by an open, loose sand substrate ( Fig. 4 View FIGURE 4 ). No data about reproduction is available, but L. cyaneinotatus is probably oviparous like L. bibronii ( Medina et al. 2008) . Data about diet came from the analysis of two guts, ant heads and beetle legs were found, with partially digested small leaves and vegetable scraps. This suggests that L. cyaneinotatus is similar to other lizards in its use of invertebrates and plants as food resources ( Videla 1983; Acosta et al. 1996; Quatrini et al. 2001; Belver & Avila 2002).
Remarks. Lizards of the Liolaemus alticolor-bibronii group have the largest latitudinal distribution in a Liolaemus species group, with L. walkeri from Junín Province in central Peru to L. bibronii in central Santa Cruz province in southern Argentina. Some species are found only in arid and semiarid environments, including Puna of Peru, Argentina, Bolivia, and Chile as L. alticolor , L. bitaeniatus , L. chaltin , L. incaicus , L. lativittatus , L. pagaburoi , L. paulinae , L. puna , L. ramirezae , L. tacnae , L. variegatus , L. walkeri , L. yanalcu , meanwhile others are restricted to some mountain ranges as L. saxatilis (Sierras de Cordoba and San Luis), L. tandiliensis (Tandil Mountains) , or particular biogeographic regions as L. araucaniensis . Liolaemus lemniscatus is found in several habitats of the western slopes of Andean mountains and in northern Andean-Patagonian forest; L. curicensis and L. fuscus are found only in central Chile region. Liolaemus gracilis is found in environments of central Argentina, but L. exploratorum is an enigmatic species described based on museum specimens and never found again in its purported range of distribution. Liolaemus bibronii is the species with probably the largest geographic range in latitude, but Morando et al. (2007) found that this may be a species complex with several candidate species.
A mtDNA gene tree analysis, including the new described species as well as other related species and candidate species of the group is depicted on Fig. 5 View FIGURE 5 . This tree is based on the mitochondrial gene fragment cyt-b (662 bp). Liolaemus cyaneinotatus is recovered within the bibronii north clade, that also includes L. gracilis , L. ramirezae , L. saxatilis , L. robertmertensi and several candidate species (Posterior Probability PP: 1). The bibronii south clade includes the haplotypes from the type locality of L. bibronii and also includes candidate species and is recovered with strong support (PP: 1). The objective of this tree is to show the position of this new species in relation with other terminal taxa of the L. bibronii complex. Species limits based on only one gene have limitations and although Liolaemus cyaneinotatus haplotypes are recovered as monophyletic, which strongly supports this new species; an integrative approach using a multi-loci species tree method and morphological analyses are needed in order to fully understand the phylogenetic relationships of the bibronii complex. A detail study with these characteristics is being carried out by our research group and will be published elsewhere.




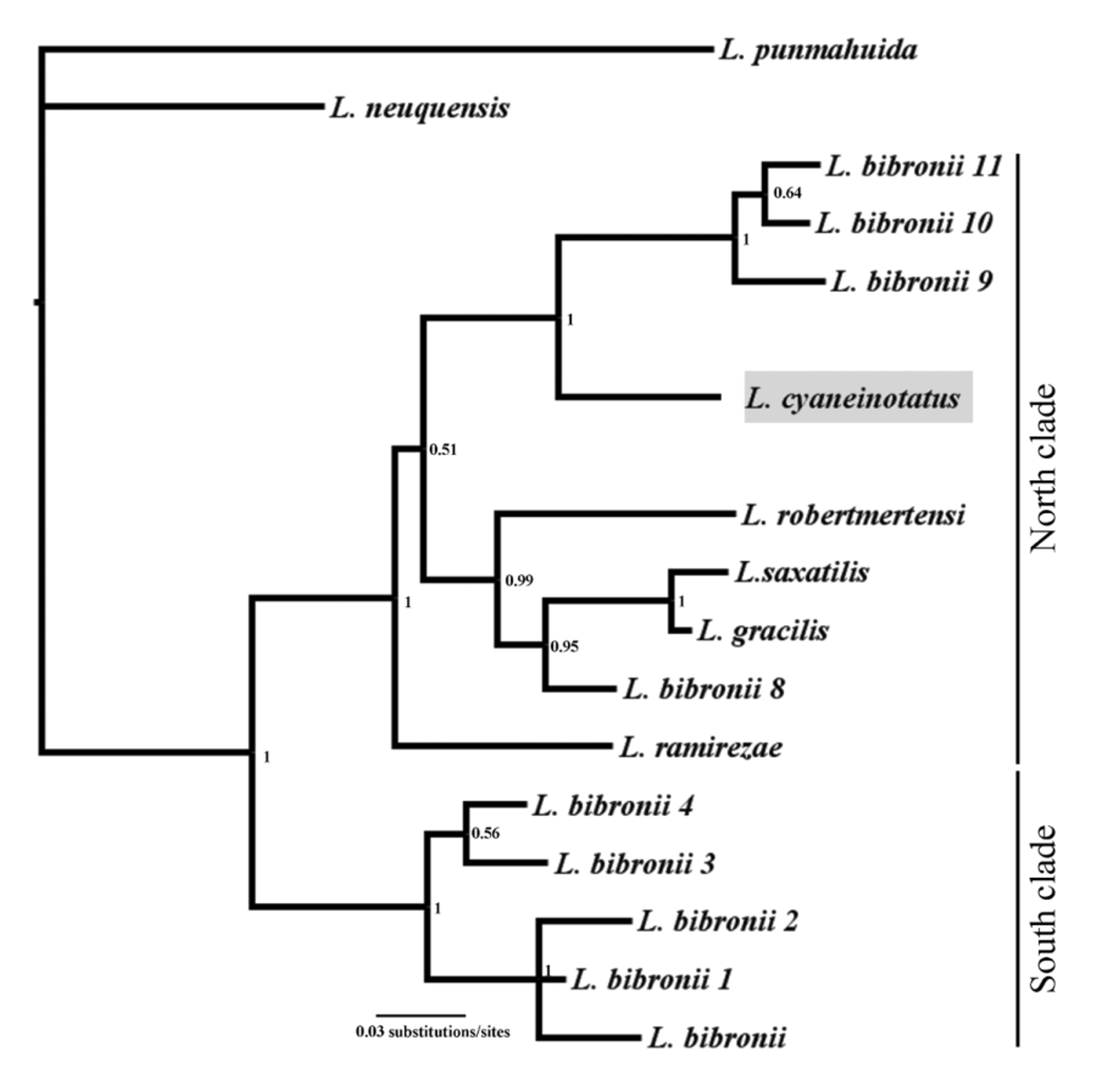
TABLE 1. Diagnostic characters among geographically proximal members of the bibronii-alticolor group; data for L. araucaniensis, L. curicencis, L. fuscus and L. lemniscatus were taken from Piñeira-Donoso and Nuñez (2005). Ranges are given in parentheses; snout-vent, axila-groin, and tail lengths in mm; and elevation range in m.
| n=15 | n=1 | n=34 | |
|---|---|---|---|
| Snout–vent length | 51.0 (37.9–59.0) | 60.9 | 60.4 (54.5–64.2) |
| Distance from axilla –groin | 21.7 (15.3–25.8) | 27.2 | 28.8 (27.0–31.7) |
| Vertebral line | present | present | irregular |
| Tail length | 82.3 (67.7–103.7) | 104.3 | 112.5 (110.0–115.0) |
| Scales around midbody | 60–68 | 58 | 49–56 |
| Infradigital lamellae of 4th toe | 21–24 | 22 | 24–27 |
| Male precloacal pores | 3–5 | 4 | 2–3 |
| Females with precloacals pores | absent | absent | absent |
| Reproductive mode | ? | oviparous | oviparous |
| Elevational range | 1300–2000 | 1400 | 1870–1950 |
TABLE 4. Between sex variation in meristic and morphometric characteristics of Liolaemus cyaneinotatus. Measurements are in mm, numbers for each variable indicate mean ± standard deviation, with range in parentheses. Precloacal pores, infra and supralabials scales are shown as ranges.
| Variable Snout–vent length | Males (n = 8) 52.4 ± 6.5(37.9–59.0) | Females (n = 7) 49.2 ± 3.9(41.6–53.7) |
|---|---|---|
| Head length Head width Distance between fore and hind limbs Humerus length | 11.5 ± 1.3(8.7–13.0) 9.2 ± 1.1(7.0–10.8) 22.8 ± 3.0(16.2–25.1) 5.9 ± 0.6(4.7–6.9) | 10.4 ± 0.6(9.4–11.3) 8.2 ± 0.7(7.1–9.0) 22.2 ± 2.5(17.7–25.8) 5.4 ± 0.3(5.1–5.9) |
| Radius-ulna length | 6.2 ± 0.9(4.6–7.7) | 5.6 ± 0.4(4.76–6.1) |
| Hand length Femur length | 8.1 ± 0.9(6.2–9.3) 7.9 ± 1.1(5.5–9.5) | 7.4 ± 0.7(6.4–8.4) 7.2 ± 0.5(6.5–7.7) |
| Tibio-fibula length Foot length | 10.5 ± 1.2(7.8–11.5) 15.4 ± 1.6(11.8–16.9) | 9.5 ± 0.5(8.6–10.2) 14.2 ± 0.7(13.1–15.2) |
| Fourth-toe length Maximum body height Maximum body width | 13.4 ± 1.4(10.2–15.1) 7.2 ± 1.0(5.3–8.6) 11.9 ± 2.2(7.9–15.5) | 12.3 ± 0.6(11.6–13.1) 7.0 ± 0.7(5.6–7.7) 12.6 ± 1.5(10.5–15.1) |
| Tail length Dorsal scales between occiput and thigh | 86.8 ± 13.3(71.8–103.7) 62.7 ± 2.0(60.0–65.0) | 79.3 ± 7.9(67.7–89.1) 61.8 ± 2.9(60–68) |
| Scales around midbody | 48.6 ± 2.2(46.0–53.0) | 50.1 ± 1.6(48–53) |
| Dorsal head scales Ventral scales Precloacal pores Scales around interparietal Enlarged supraoculars | 12.1 ± 0.3(12.0–13.0) 83.1 ± 2.3(80.0–86.0) 3–5 6.6 ± 1.0(6–9) 4.4 ± 0.7(4–6) | 12 84.4 ± 6.1(75–91) 0 6.0 ± 0.6(5–7) 4.8 ± 0.9(4–6) |
| Number of temporals Number of neck scales Number of gulars Enlarged supralabials | 7.4 ± 0.5(7–8) 14.9 ± 1.5(13–17) 28.4 ± 1.6(27–31) 5–6 | 7.1 ± 0.9(6–9) 14.4 ± 1.1(13–16) 29.0 ± 2.7(24–32) 6–7 |
| Enlarged infralabials | 4–5 | 4 |
| Scales around nasal Scales between rostral and frontal | 6.7 ± 0.4(6–7) 5.2 ± 0.4(5–6) | 7 5.6 ± 0.8(5–7) |
| Scales between frontal and supercilliaries Loreolabials scales | 4.6 ± 0.7(3–5) 5.9 ± 0.6 (5–7) | 4.1 ± 0.4(4–5) 5.3 ± 0.5(5–6) |
| Infradigital lamellae of 4th toe of the hand Infradigital lamellae of 4th toe of the pes | 17.5 ± 1.4(15–19) 23.5 ± 0.9(22–24) | 16.6 ± 0.5(16–17) 22.6 ± 1.1(21–24) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.