
Magelona nanseni, Mortimer & Kongsrud & Willassen, 2022
|
publication ID |
https://doi.org/ 10.1093/zoolinnean/zlab070 |
|
publication LSID |
lsid:zoobank.org:pub:278AA1B0-674E-414D-A47A-D87F43E2D6E4 |
|
DOI |
https://doi.org/10.5281/zenodo.6459434 |
|
persistent identifier |
https://treatment.plazi.org/id/039087DB-FFEA-FFF8-4243-74FBFAFEFBC4 |
|
treatment provided by |
Plazi |
|
scientific name |
Magelona nanseni |
| status |
sp. nov. |
MAGELONA NANSENI View in CoL SP. NOV.
( FIGS 11 View Figure 11 , 12 View Figure 12 )
Z o o b a n k r e g i s t r a t i o n: u r n: l s i d: z o o b a n k. org:act: 6F8FA46E-BEBB-48E0-8758-26750FCDA54E.
Type locality: Nigeria: 4.1482°N 5.7688°E, 25 m depth GoogleMaps .
Type material: Holotype, Nigeria: St. 5N–11, af in 75% Etoh ( ZMBN132141 View Materials ) . Paratypes: Liberia: St. 7LI–01, 1af in 75% Etoh ( ZMBN132142 View Materials ) ; St. 7LI–04, 1af in 75%Etoh (NMW.Z.2021.001.0006); Ghana: St. 2009105–GP1/28, 1af in 75% Etoh ( ZMBN107275 View Materials ) ; St. 2009105–GE1/28, 5af in 75% Etoh ( ZMBN107276 View Materials ) . Nigeria: St. 5N–11, 1af in 96% Etoh ( ZMBN107336 View Materials , DNA-voucher) .
Etymology: The new species is named in honour of the zoologist and oceanographer Dr Fridtjof Nansen, whose name is also linked with the research vessel from which all samples from West Africa included in this paper were collected.
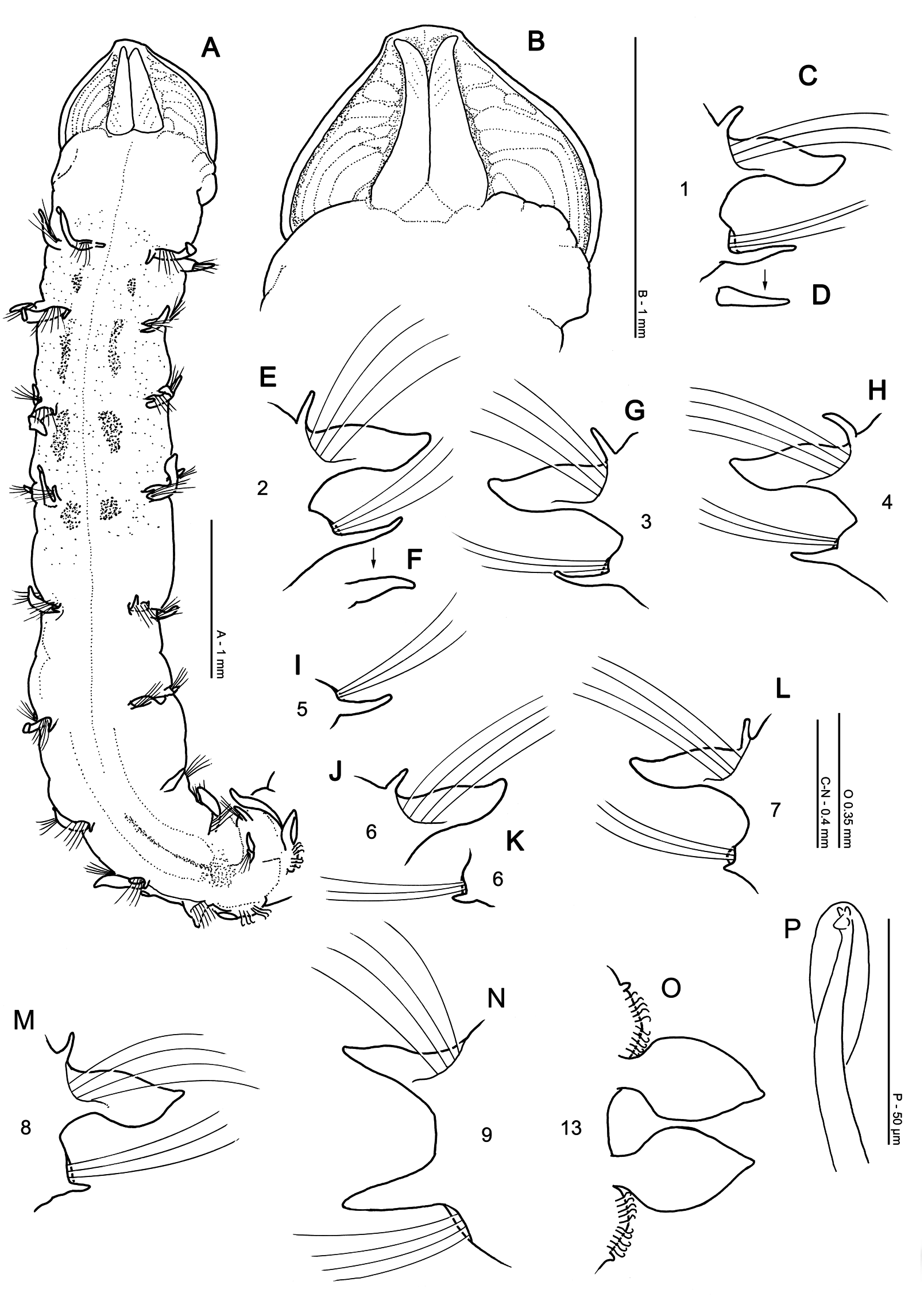
Diagnosis: Prostomium wider than long, with rudimentary prostomial horns. Chaetigers 1–8 with slender foliaceous notopodial lamellae, with long, superior dorsal lobes and slender, triangular neuropodial lamellae. Lamellae of chaetiger 9 triangular postchaetal in both rami. All thoracic chaetae capillary. Abdominal lateral lamellae basally constricted and spatulate, with pointed tips. Abdominal hooded hooks tridentate, in two groups, vis-à-vis. No pouches observed, pygidium unknown.
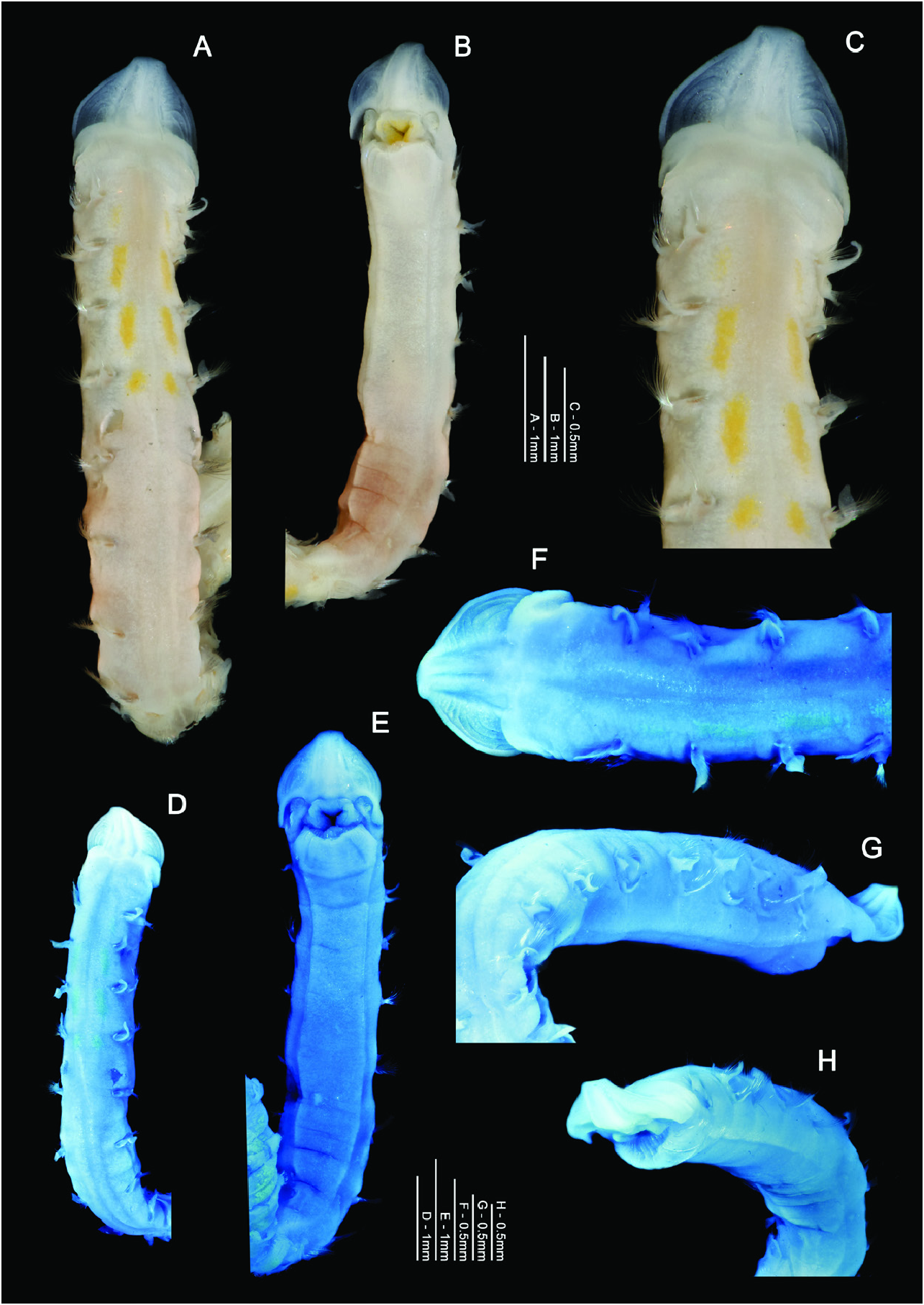
Description: A stout species; junction between thorax and abdomen noticeable, although not distinctly marked ( Figs 11A View Figure 11 , 12B View Figure 12 ), abdomen more rounded than thorax. Holotype, anterior fragment: prostomium 1.0 mm wide, 0.85 mm long; thorax 5.75 mm long (including prostomium), 1.1 mm wide (at its widest point, approximately chaetiger 6); abdomen 0.9 mm wide; total length approximately 13.25 mm for 26 chaetigers. (NB last chaetiger of holotype dissected and slide-mounted. Width measurements not including parapodia.) Thorax tapering towards abdomen from approximately chaetiger 7. Additional anterior fragment (ZMBN107336) 4.8 mm long for 11 chaetigers, approximately 0.8 mm wide. Other anterior fragments 5.5–11.5 mm for 14–20 chaetigers.
Prostomium rounded laterally ( Figs 11A, B View Figure 11 , 12A–F View Figure 12 ), wider than long (L: W ratio 0.85). Proximal prostomial corners extend around bases of palps towards ventral surface ( Fig. 12B, E, H View Figure 12 ). Distally, prostomium squared into ‘rudimentary horns’, anterior margin roughly straight but with a small, medial rise on holotype. One pair of distinct, longitudinal, dorsal muscular prostomial ridges, thick and abutting for majority of length, although diverging for distal quarter. No obvious outer pair of ridges. Distinct arched markings either side of ridges, becoming smaller transverse markings towards distal tip. Small, white speckles present sporadically on surface of prostomial markings. Burrowing organ partially everted in one specimen (ZMBN107336), oval with light longitudinal stripes inferiorly. Buccal region on ventral side of prostomium comprised of three lips: a larger triangular upper lip, surmounting two smaller lateral lips ( Fig. 12B, E View Figure 12 ), inner surface of which is heavily speckled, cream/yellow in colour. Palps retained on one specimen (ZMBN107336, RH palp now detached but present in vial), thick, short and appearing frilly due to length and abundance of papillae. Palps approximately 7 mm long, non-papillated region reaching approximately chaetiger 3. Papillae long, proximally five rows either side of an inconspicuous mid-palp line, devoid of papillae, four rows either side medially and two rows distally. Only palp stumps either side of buccal region retained on holotype ( Fig. 12E View Figure 12 ).
Achaetous region behind the prostomium, roughly twice the length of chaetiger 1 ( Figs 11A View Figure 11 , 12A, C, F View Figure 12 ). Chaetigers 1–8 similar; parapodia biramous ( Fig. 11C–M View Figure 11 ). Notopodia with low, rounded, prechaetal lamellae confluent with slender, smooth-edged, foliaceous, postchaetal lamellae with rounded tips, of similar size along thorax. Those of chaetiger 8 slightly more foliaceous, with pointier tips. Single, long, tapering, superior dorsal lobe present on all thoracic chaetigers, except chaetiger 9, decreasing slightly in size in posterior thorax. Neuropodia with low pre- and postchaetal lamellae, encircling chaetae cufflike and confluent with slender, triangular lamellae, directly under chaetae ( Fig. 11D, F View Figure 11 ), decreasing in size in posterior thorax. Initially slightly prechaetal but becoming completely ventral by chaetiger 4.
Chaetiger 9: shorter and narrower than preceding chaetigers ( Figs 11A View Figure 11 , 12B View Figure 12 ). Notopodial prechaetal lamellae low, confluent with larger, slender, triangular, postchaetal lamellae, of similar length to preceding chaetigers ( Fig. 11N View Figure 11 ). No superior dorsal lobes observed. Neuropodia similar, but chaetae emerging below postchaetal lamellae from low inferior ridge. Occasionally inferior edge of ridge slightly raised but never appearing as a distinct process. Chaetae of chaetigers 1–9 simple bilimbate, winged capillaries.
Parapodia of abdominal chaetigers ( Fig. 11O View Figure 11 ) with broad, spatulate, lateral lamellae with pointed tips, of similar size in both rami. Basally constricted, but with no obvious postchaetal expansion behind chaetal rows. Small, triangular processes (DML, VML) observed at inner margins of chaetal rows. Abdominal chaetae tridentate hooded hooks ( Fig. 11P View Figure 11 ) of a similar size, superior two fangs parallel, above main fang. Hooks in two approximately equal groups for each ramus, main fangs vis-à-vis ( Fig. 11O View Figure 11 ). Approximately ten hooks per ramus in the anterior abdomen. No abdominal pouches observed, but material only possessing up to 26 abdominal chaetigers. Pygidium unknown.
Colour: No living material observed. Preserved specimens cream in colour, faint reddish to brown pigment present in the posterior thorax, between chaetigers 5–8 ( Fig. 12A, B View Figure 12 ). Pigmentation slightly darker in 96 % ethanol preserved specimen (ZMBN107336), particularly ventrally. Pigmentation of freshly preserved specimens unknown, pigment is likely to have faded to some degree for the holotype after 13 years of preservation. Obvious elongate, yellow, dorsal patches (glandular?) present between chaetigers 1–5, adjacent to parapodia and either side of the middorsal line ( Fig. 12A, C View Figure 12 ). Staining with methyl green ( Fig. 12D–H View Figure 12 ) shows no distinct pattern, just weak stain all over. However, speckled areas of the prostomium, thoracic region, as well as interparapodial abdominal patches, more noticeable in stained specimens.
Distribution: Only known from five stations, off three countries, Liberia to Nigeria, at depths of 25–29 m ( Fig. 1 View Figure 1 ).
Remarks: This new species can be distinguished from all other pigmented magelonid species within the MIWA region based on the following characters. In possessing thoracic superior dorsal lobes, M. nanseni differs from M. alleni and M. fasciata , in which they are absent. In possessing tridentate abdominal hooks, it further differs from M. fasciata and, additionally, M. mackiei , in which they are bidentate. It also differs from the latter two species and M. picta in not having small processes below the chaetal bundles in the neuropodia of chaetiger 9. Of the MIWA species, M. nanseni is closest to M. picta , as noted above, but differs in having less foliaceous notopodial lamellae in the thorax, more basally constricted abdominal lamellae and in terms of body pigmentation (see above).
Of the other magelonid species known to carry posterior thoracic pigmentation, M. nanseni differs from M. cincta , M. equilamellae , M. japonica , M. symmetrica , M. polydentata and M. variolamellata in possessing thoracic superior dorsal lobes (absent in the others). It further differs from M. polydentata in possessing tridentate hooded hooks in the abdomen rather than polydentate.
Of all the previously described African species, M. nanseni shares affinities with M. cepiceps from the Seychelles, particularly in the nature of the prostomium, which is somewhat onion-shaped in the latter species. However, they differ in the lamellae of chaetiger 9; the latter species possessing thoracic superior dorsal lobes on all thoracic chaetigers (absent on chaetiger 9 in M. nanseni ) and low, rounded, postchaetal neuropodial lamellae, with small, ventral processes below the chaetal bundle (as opposed to slender, elongate triangular postchaetal lamellae for the new species).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.