
Melayonchis aileenae Dayrat and Goulding, 2017
|
publication ID |
https://doi.org/ 10.1080/00222933.2017.1347297 |
|
publication LSID |
lsid:zoobank.org:pub:671922DB-C6C1-44A5-B2CD-A3A3127CB668 |
|
persistent identifier |
https://treatment.plazi.org/id/A496D3C9-D454-4B32-9A84-C732514F7554 |
|
taxon LSID |
lsid:zoobank.org:act:A496D3C9-D454-4B32-9A84-C732514F7554 |
|
treatment provided by |
Felipe |
|
scientific name |
Melayonchis aileenae Dayrat and Goulding |
| status |
sp. nov. |
Melayonchis aileenae Dayrat and Goulding View in CoL sp. nov.
( Figures 20–26 View Figure 20 View Figure 21 View Figure 22 View Figure 23 View Figure 24 View Figure 25 View Figure 26 )
Type locality
Malaysia, Peninsular Malaysia, Kuala Sepatang, 04°50.434 N, 100°38.176 E, 18 July 2011 [station 27, old forest with tall, old Rhizophora trees, high in the tidal zone (ferns), following boardwalk in educational preserve, reached a creek lower in the tidal zone, with mud] ( USMMC 00018 ) GoogleMaps .
Type material
Holotype, designated here: one specimen 20/10 [DNA 970] mm, in 95% alcohol, leg. B. Dayrat and T. Goulding ( USMMC 00018 ). Note that other specimens (not part of the type material) were collected from the same locality and are mentioned below in the additional material examined.
Additional material examined
Malaysia, Peninsular Malaysia, Kuala Sepatang, 04°50.434 N, 100°38.176 E, 18 July 2011, 8 specimens (25/16 [#1] to 15/ 8 mm; 20/12 [#2]), leg. B. Dayrat and T. Goulding [station 27, old forest with old, tall Rhizophora trees, high in the tidal zone (ferns), following boardwalk in educational preserve, reached a creek lower in the tidal zone, with mud] ( USMMC 00019); Malaysia, Peninsular Malaysia, Matang, off Kuala Sepatang, Crocodile River, Sungai Babi Manpus, 04°49.097 ʹ N, 100°37.370 ʹ E, 19 July 2011, 1 specimen (18/14 [DNA 968] mm), leg. B. Dayrat and T. Goulding [station 28, old and open Rhizophora forest with tall trees, hard mud, creeks, and many dead logs] ( USMMC 00020); Malaysia, Peninsular Malaysia, Matang, close to the jetty, facing the fishermen’ s village on the other side of the river, 04°50.154 ʹ N, 100°36.368 ʹ E, 20 July 2011, 1 specimen (25/ 10 mm), leg. B. Dayrat and T. Goulding [station 29, oldest and open Rhizophora forest of tallest and beautiful trees, with hard mud, many creeks, and many dead logs] ( USMMC 00021); Brunei Darussalam, Mentiri, Jalan Batu Marang, 04°59.131 ʹ N, 115°01.820 ʹ E, 29 July 2011, 1 specimen (13/8 [DNA 1047] mm), leg. B. Dayrat, T. Goulding and S. Calloway [station 34, old mangrove with tall Rhizophora trees with high roots and Thalassina mounds] ( BDMNH); Vietnam, Can Gio, 10°24.108 ʹ N, 106°53.019 ʹ E, 13 July 2015, 2 specimens (19/12 [DNA 5606] and 15/ 12 mm), leg. T. and J. Goulding [station 224, mangrove with big Rhizophora trees mixed with Avicennia ] ( ITBZC IM 00005); Vietnam, Can Gio, 10°27.803 ʹ N, 106°53.288 ʹ E, 16 July 2015, 3 specimens (25/11 to 14/ 10 mm), leg. T. and J. Goulding [station 228, high intertidal, open forest of tall Rhizophora trees, hard mud, near a small river] ( ITBZC IM 00006); Vietnam, Can Gio, 10°27.620 ʹ N, 106°53.316 ʹ E, 17 July 2015, 1 specimen (20/10 [DNA 5631] mm), leg. T. and J. Goulding [station 231, open mangrove with large Avicennia trees, soft mud, some dead logs] ( ITBZC IM 00007); Vietnam, Can Gio, 10°27.692 ʹ N, 106°53.308 ʹ E, 18 July 2015, 5 specimens (32/20 to 20/ 14 mm), leg. T. and J. Goulding [station 232, high intertidal, Avicennia mangrove separated from tall Rhizophora trees (station 228) by a road] ( ITBZC IM 00008). India, Andaman Islands, Middle Andaman, Rangat, Shyamkund, 12°28.953 ʹ N, 092°50.638 ʹ E, 11 January 2011, 1 specimen 14/ 8 mm, leg. B. Dayrat and V. Bhave [station 59, by a large river, deep mangrove with tall trees, small creeks, and plenty of dead muddy logs, next to a road and a small cemented bridge above a creek] ( BNHS 90).
Distribution
Malaysia (type locality), India ( Andaman Islands ), Brunei Darussalam and Vietnam .
Etymology
Melayonchis aileenae is dedicated to our dear friend Dr Tan Shau Hwai (Aileen), professor at the Universiti Sains Malaysia, Penang, whose generous help was critical for us to conduct field work in Malaysia and other parts of South East Asia.
Habitat ( Figure 20 View Figure 20 )
Melayonchis aileenae is found in the high intertidal and it especially favours old Rhizophora forests. It lives on trunks and roots of mangrove trees, often not muddy but covered with algae instead. It can also be found on dead logs. In Vietnam, a few specimens were found under rotting bark on mangrove tree roots. Occasionally, it can also be found on cemented walls at the margin of a mangrove. It is not found directly on mud.
Abundance
Melayonchis aileenae is a rare species. It was found at only a few of the dozens of mangrove sites that we visited in the region, and we found only a few specimens. It is not particularly cryptic so we believe that it actually is uncommon.
Colour and morphology of live animals ( Figures 21 View Figure 21 , 22 View Figure 22 )
Live animals are usually not covered dorsally with a thin layer of muddy mucus and the colour of their dorsal notum can readily be seen. Occasionally, live animals are covered with a thin layer of mud which first needs to be removed to see the dorsal colour. The background of the dorsal notum varies from light brown to black. The notum bears additional markings (white dots and inconspicuous darker, irregular, longitudinal lines) which vary among individuals. The colour of the hyponotum is very light and varies from cream to light grey. Because it is lightly coloured, its margin is not marked by a significantly lighter ring. However, there is a ring of white dots around the margin of the hyponotum that could correspond to gland openings. The foot is cream to pale yellow ( Figures 21e, f View Figure 21 , 22f View Figure 22 ). The brown ocular tentacles are short and extend for only a few millimetres beyond the notum margin when the animal crawls undisturbed. The head is small and remains covered by the dorsal notum as the animal crawls.
The body is not flattened. The dorsal notum is elongated, oval. The dorsal notum is not particularly thick. Its surface is remarkably smooth (occasionally very finely granular). Dorsal gills are absent. Large or small papillae are absent. ‘Dorsal eyes’ are present and at the tip of very tiny tubercles which can be barely noticeable on the notum surface. There are about 20 isolated ‘dorsal eyes’ throughout the dorsal notum. In the centre, there are three ‘dorsal eyes’ clustered together. When disturbed, animals do not coil up into a sphere. Crawling individuals can measure up to 32 mm, but most of them measure about 20 mm on average.
External morphology
Some of the distinct colour traits of live animals are lost in preservation (and this clearly worsens with time). The dorsal colour of preserved animals varies from homogeneously cream to black with occasional, irregular, longitudinal lines. The preserved foot tends to be slightly yellow while the hyponotum is white. The width of the hyponotum relative to the width of the pedal sole varies among individuals but represents on average about 1/ 3 of the total width ( Figures 21e, f View Figure 21 , 22f View Figure 22 ).
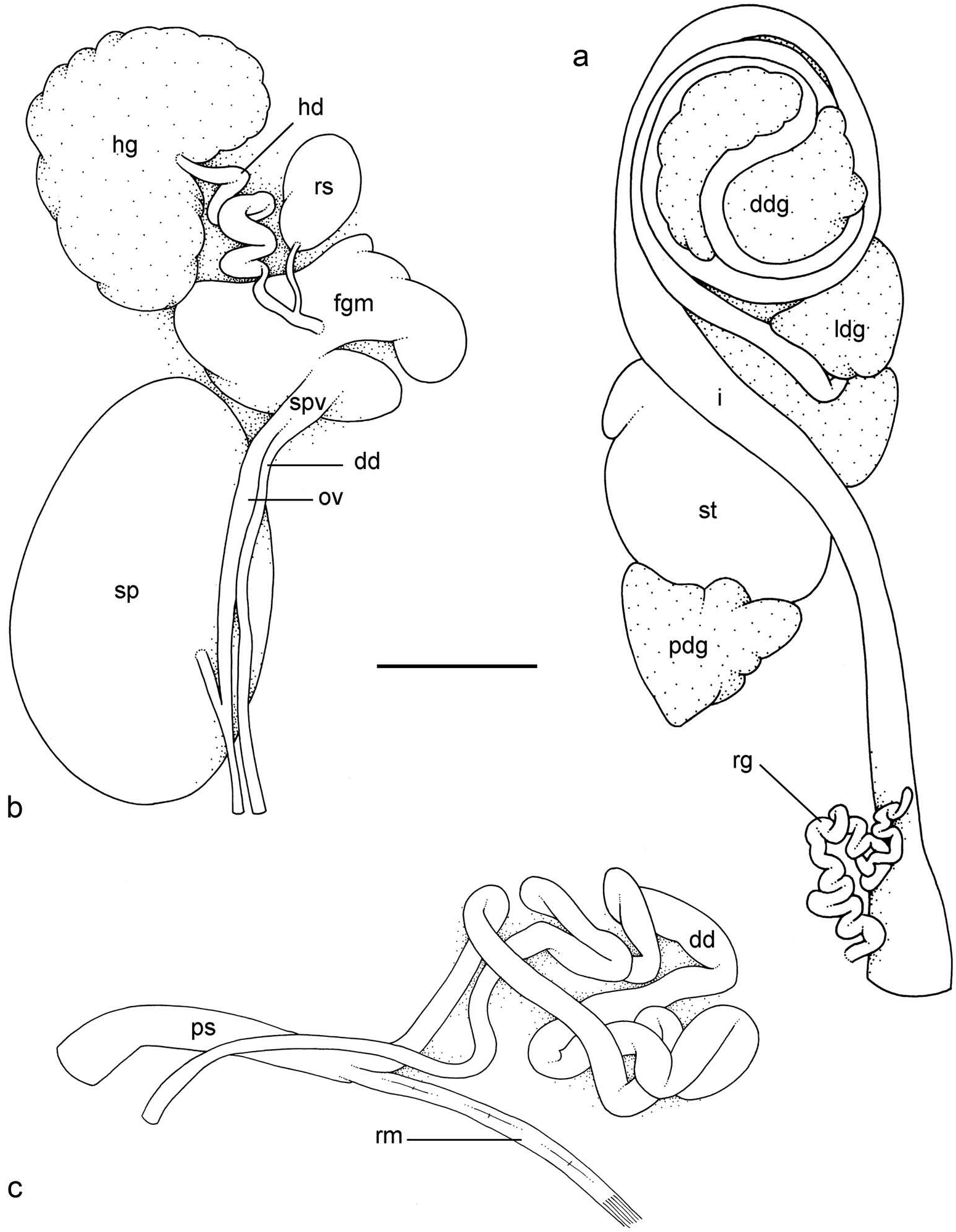
Digestive system ( Figures 23a View Figure 23 , 24 View Figure 24 , 25 View Figure 25 )
Examples of radular formulae are ( Figures 24 View Figure 24 , 25 View Figure 25 ): 100 × (290–1–290) in USMMC 00019 #1 (25 mm long), 140 × (380–1–380) in USMMC 00019 #2 (20 mm long), 95 × (250–1–250) in USMMC 00021 (25 mm long), and 95 × (250–1–250) in ITBZC IM 00007 (20 mm long). Along a half row, all the lateral teeth do not have exactly the same length and shape. The length of the hook of the lateral teeth gradually increases (from innermost to outermost) from about 25–35 µm to about 40–50 µm, excluding the first few (about 5) innermost and outermost lateral teeth which are significantly smaller than the rest of the lateral teeth. Finally, the tip of the hook of the lateral teeth is quite variable and can be pointed, tapered or round. The pattern of its loops is intermediary between types II and III ( Figure 23a View Figure 23 ).
Reproductive system ( Figures 23b, c View Figure 23 , 26 View Figure 26 )
There is a small, pear-shaped, slightly bent receptaculum seminalis (caecum) along the hermaphroditic duct. The oval spermatheca (for the storage of exosperm) is very large and connects to the oviduct through a very short duct ( Figure 23b View Figure 23 ).
The male anterior organs consist of the penial complex (penis, penial sheath, vestibule, deferent duct, retractor muscle) ( Figure 23c View Figure 23 ). There is no penial accessory gland. The penial sheath is short (less than 2 mm in length), straight and of similar width to the distal vestibule. The penial sheath protects a penis which consists of a short, slightly conical papilla of about 0.5 mm long ( Figure 26 View Figure 26 ). The penial papilla is slightly longer and more elongated (0.8 mm) in Vietnam ( Figure 26a View Figure 26 ). There are no penial hooks. The vestibule is continuous with and similar to the penial sheath. The insertion of the retractor muscle marks the separation between the penial sheath and the deferent duct. The retractor muscle is short (as long as the penial sheath) and inserts in the anterior part of the visceral cavity. The deferent duct is highly convoluted with many loops, even though it is significantly less convoluted in immature specimens ( Figure 23c View Figure 23 ).
Distinctive field diagnostic features
A table at the end of the introduction summarises the most important features that can help distinguish and identify Melayonchis species ( Table 3). In the field, M. aileenae is easy to identify because no other species looks like it. Its dorsal colour (brown background with lighter and darker markings), although variable, combined with its exceptionally smooth notum, make it unique and unmistakable.






| T |
Tavera, Department of Geology and Geophysics |
| IM |
Indian Museum |
| V |
Royal British Columbia Museum - Herbarium |
| BNHS |
Bombay Natural History Society |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.