
Myxicola steenstrupi Krøyer, 1856
|
publication ID |
https://doi.org/ 10.5852/ejt.2023.900.2305 |
|
publication LSID |
lsid:zoobank.org:pub:964E1F58-F939-47FC-B285-2953D61A442F |
|
DOI |
https://doi.org/10.5281/zenodo.10164562 |
|
persistent identifier |
https://treatment.plazi.org/id/03B80677-FFEF-FF3E-FD80-BB84685FF92C |
|
treatment provided by |
Plazi |
|
scientific name |
Myxicola steenstrupi Krøyer, 1856 |
| status |
|
Myxicola steenstrupi Krøyer, 1856 View in CoL
Material examined
Syntypes FAROE ISLANDS • 1 spec.; Tórshavn; NHMD 109911 .
GREENLAND • 1 spec.; NHMD 109912 .
Additional material
FAROE ISLANDS • 1 spec., 1 juv.; head of Kaldbakfjord; depth 1–2 m; P. Knight-Jones leg.; from abandoned Mytilus ropes; NMW.Z.2009.038.0673 .
Description
Faroese specimens
Faroe Islands syntype complete, in 2 pieces, with 54 chaetigers, 7–8 thoracic (difficult to determine due to condition) and 46 abdominal; body length 26 mm, crown length 11 mm. Other specimens complete, with 46 and 50 chaetigers, of which 8 are thoracic; body lengths 12 and 31 mm, crown lengths 5 and 9 mm. Preserved colour cream (NMW.Z) or pale brown (syntype), no pigment observed. 1–2 minute interramal (thoracic) and lateral (abdominal) eyespots present on NMW.Z specimens only, as well as pygidial eyespots, pygidium tip missing on type.
Radiolar crown with semicircular radiolar lobes bearing 20 pairs of radioles (type) or 12 and 17 pairs, respectively, on juvenile and larger NMW.Z specimens. Connecting basal membrane long (damaged between most radioles on type). All radiole tips damaged except on juvenile: very high basal membrane with free portion of tip less than 10% total length of radiole, tip tapering to a blunt point; pinnulae long, thin, ca 0.25 × total length of radiole, terminating below lower margin of basal membrane. Radiolar eyes absent. Dorsal lips with small, digitiform radiolar appendages, arising centrally, surrounded by enlarged, lobate ventral lips that overlap centrally, connected to radiolar lobes dorsally; pinnular appendages absent; ventral radiolar appendages absent; parallel lamellae and ventral sacs both absent. Anterior peristomial ring with small ventral, triangular lobe, concave. Lateral notches present. Notopodia as oval cushions with fine notochaetae, number difficult to assess due to damage, but less than 100. Form of notochaetae and uncini not observed. Tube not present.
Greenlandic syntype
Syntype complete with 55 chaetigers, 8 thoracic and 47 abdominal; body length 41 mm, crown length 15 mm. Preserved colour pale brown, no pigment or eyespots observed. Body shape unclear due to extent of dissection, but widest anteriorly, tapering to pygidium, tip missing. Radiolar crown with semicircular radiolar lobes bearing 20–21 pairs of radioles. Radioles with wide radiolar flanges and long, tapered tips, approximately 23% of total radiole length. Pinnulae very fine, dense, longest in distal part of radiole, up to 0.25 × total length of radiole, then rapidly decreasing in length, terminating at or above lower margin of basal membrane. Radiolar eyes absent. Dorsal lips with slender, digitiform radiolar appendages, arising centrally, surrounded by enlarged, lobate ventral lips connected to radiolar lobes dorsally; pinnular appendages absent; ventral radiolar appendages absent; parallel lamellae and ventral sacs both absent. Anterior peristomial ring too damaged and degraded to describe. Notopodia as oval cushions, notochaetae all broken. Uncini not observed. Pygidial tip missing, tube not present.
Remarks
The two type specimens from the Faroes and Greenland were borrowed from the Natural History Museum of Denmark and examined along with two additional specimens in NMW.Z from the Faroe Islands collected by Phyllis Knight-Jones. Both type specimens were in poor condition and many of those characters that have been detailed above for M. infundibulum and M. polychroma sp. nov. could not be observed. The larger NMW.Z specimen also had a badly damaged crown, limiting observations on the crown and dorsal lips, and the remaining specimen was a juvenile, although in good condition. No attempt is therefore made here to re-validate the species. Although it is considered that the species is likely to be found to be valid in the future, to do so will require the examination and/or collection of new specimens from the type localities to gather the necessary morphological details and, ideally, tissue samples for comparative genetic work. In the meantime, the details above are noted.
Both syntypes had labels with remarks from Mary Petersen stating that they were specimens of Krøyer’s, listed in his unpublished catalogue of worms received by NHMD from the Royal Museum, and in that way they had been identified as syntypes for the described species. The two syntypes are very different in size, the type from Greenland being nearly twice as large as that from Faroe Islands, but for roughly the same number of chaetigers and radioles. The NMW.Z specimens were of a similar size to the Faroe Island type, with similar numbers of chaetigers and radioles. The damage and degradation on both types made some observations on the body and chaetae almost impossible as well as on the crown of the Faroe Islands type. The crown of the Greenland type was in a much better condition. The two types show different characteristics in the radioles and dorsal lips, leading to the conclusion that they represent different taxa, although further observations on the chaetae are also needed. This also presents a dilemma as to which type specimen would be designated for the name. Although it is believed that Myxicola steenstrupi is likely to be recognised as a valid species in the future, at the present time it must wait until new specimens are available and a detailed description can be developed in conjunction with one of the available type specimens.
DNA Results
The UK Myxicola infundibulum specimens showed no genetic variation within the 16S and COI genes, with only a single haplotype being present for each. The same 16S haplotype was also retrieved from all Australian specimens, except one, and the specimen from Croatia, both of which showed small variations (16S: 0.0042 and 0.0085) from the rest. For COI, fewer Australian sequences were available, and none for the Mediterranean, but of the six that were available, all were identical to the UK specimens except for the one from Adelaide (K2P = 0.0109). Of the United States and Canadian M. ‘ infundibulum ’ and Myxicola sp. sequences analysed, either from GenBank or from Dane (2008), none returned a K2P distance less than 0.2154 (COI) or 0.0389 (16S) from the ‘true’ M. infundibulum . The COI sequences for Myxicola aesthetica , on the other hand, were closer to those for M. ‘ infundibulum ’ from the USA and Canada (K2P = 0.1214 –0.1768) than they were to any UK or Australian specimens (K2P = 0.2154 –0.2559) of M. infundibulum or M. polychroma sp. nov. Myxicola polychroma sp. nov. showed a similar scale of distance from ‘true’ M. infundibulum as it did from United States and Canadian M. ‘ infundibulum ’.
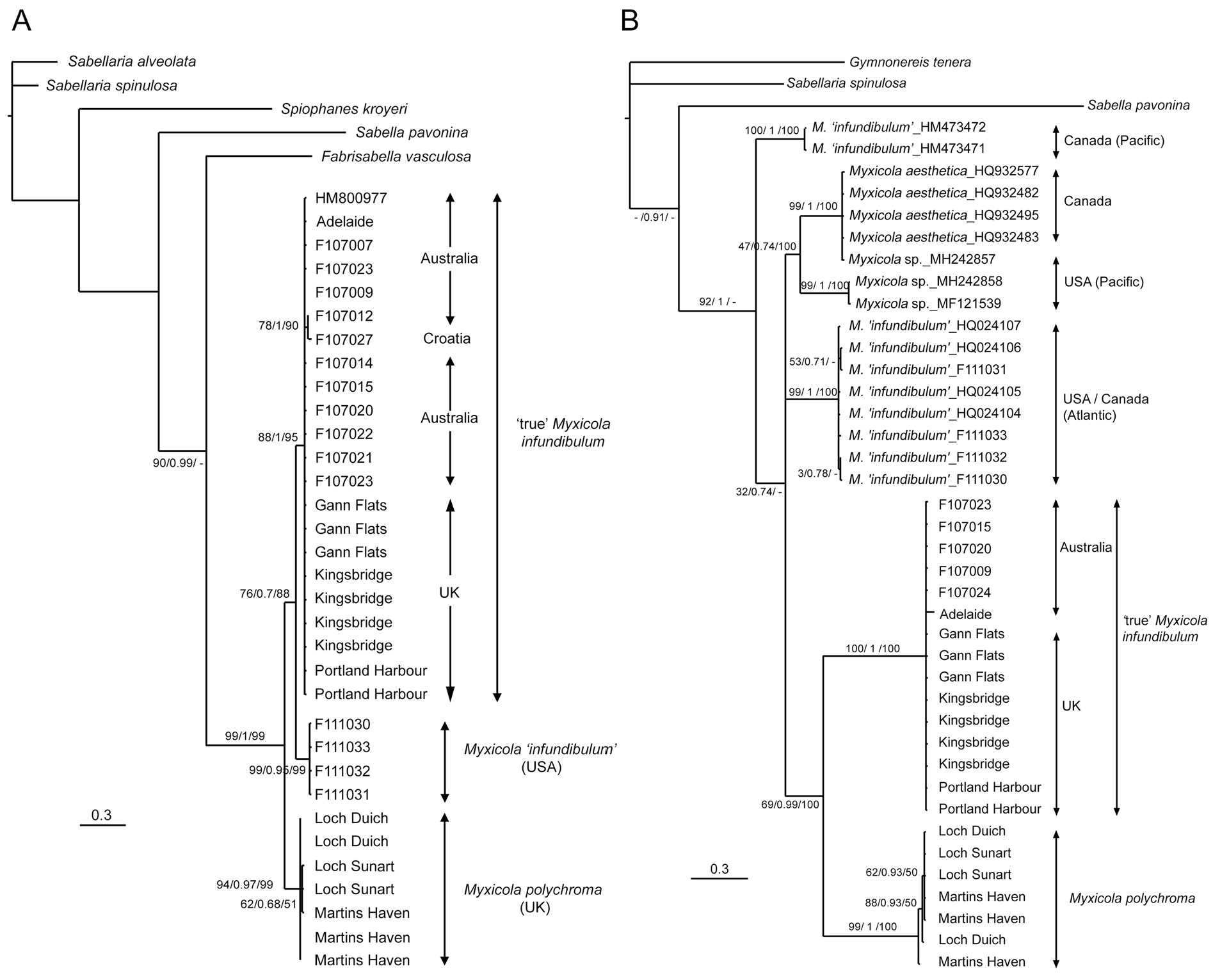
Maximum parsimony returned 7 most parsimonius trees for the 16S dataset (tree length: 477; CI = 0.7862). All trees contained the same major clades (outgroups, ‘true’ infundibulum , USA Myxicola and M. polychroma sp. nov.), only differing in the relationships within the ‘true’ infundibulum clade, all of which had little support. Two most parsimonius trees were returned for the COI dataset (tree length = 651; CI = 0.6559), the main difference between them being the position of the M. polychroma sp. nov. clade with respect to the outgroups and the other taxa of Myxicola . Maximum likelihood, Bayesian inference and the maximum parsimony consensus trees for both genes all returned similar tree topologies with the same well-supported overarching clades ( Fig. 11A–B View Fig ).
In both the 16S and COI datasets, those specimens identified as Myxicola infundibulum from the UK, including those from the type locality, formed a strongly supported distinct clade along with all specimens from Australia and, for 16S, the specimen from Croatia (16S: 88–95% bootstrap support, 0.7 Bayesian posterior probability; COI: 100% bootstrap support, 1.0 posterior probability). There was no distinction between the populations around the UK or between the UK, Australia and Croatia ( Fig. 11A View Fig ). Myxicola aesthetica formed a separate clade, along with one of the Myxicola sp. sequences, in the COI tree, indicating a probable identity match ( Fig. 11B View Fig ). The remaining two Myxicola sp. sequences formed their own well-supported clade, sister to that of Myxicola aesthetica , and it is likely that they represent an as-yet undescribed M. aesthetica -like taxon. All USA (16S) and USA and Canadian (COI) M. ‘ infundibulum ’ sequences formed a strongly-supported distinct clade for both genes ( Fig. 11A–B View Fig ), confirming their different identity and the restriction of M. infundibulum ’s native range to the northeast Atlantic, except where anthropogenically introduced.
Myxicola polychroma sp. nov. formed a strongly or fully supported clade in all analyses, although its affinities to the other taxa were not completely resolved, with the COI tree returning it as sister to ‘true’ infundibulum ( Fig. 11B View Fig ), but the 16S analyses finding it to be sister to a combined M. infundibulum and USA Myxicola clade ( Fig. 11A View Fig ). The latter analyses, however, did not include any M. aesthetica or Pacific Myxicola sequences that would provide a wider comparison of taxa.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.