
Neosclerocalyptus gouldi, Zurita & Carlini & Scillato-Yane, 2008
|
publication ID |
https://doi.org/ 10.5281/zenodo.4651202 |
|
persistent identifier |
https://treatment.plazi.org/id/573A87B2-FFF7-454B-E6AC-FA66FDA34969 |
|
treatment provided by |
Felipe |
|
scientific name |
Neosclerocalyptus gouldi |
| status |
sp. nov. |
Neosclerocalyptus gouldi n. sp. ( Fig. 1 View FIG )
HOLOTYPE. — MCA 2010 View Materials , skull and cephalic shield, right humerus and fragmentary dorsal carapace.
ETYMOLOGY. — Named in honour of Stephen Jay Gould (1941-2002), outstanding paleontologist and popular
science writer, who was also a fierce critic of all forms
of oppression and of the social use of science as an ideological foundation for power.
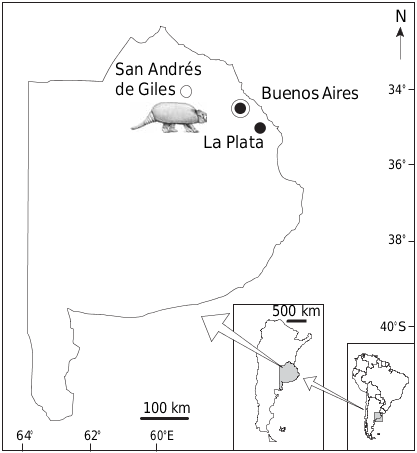
TYPE LOCALITY. — Intersection of Route no. 41 and earth road to the town of San Andrés de Giles, Buenos Aires, Argentina (34°26’57”S, 59°26’56”W) ( Fig. 2 View FIG ).
CHRONOLOGICAL AND GEOGRAPHICAL DISTRIBUTION. — Buenos Aires Formation. Middle Pleistocene (Bonaerian Stage; c. 0.40-0.13 Ma). San Andrés de Giles , Buenos Aires Province, Argentina .
DIAGNOSIS. — A Hoplophorini similar to N. paskoensis ( Zurita, 2002) but more robust, with a narrower skull and more extended anteroposteriorly (MDZA/LE: 0.66) ( Table 1). The dorsal profile of skull is convex, with the parieto-occipital region directed downwards as in N. paskoensis and Eosclerocalyptus tapinocephalus
(Cabrera, 1939), although less markedly so than in these
A new species of Neosclerocalyptus ( Xenarthra , Glyptodontidae ) from the Pleistocene of Argentina
D, ventral view; F, occlusal view. Scale bar: 1 cm. Zurita A. E. et al.
species. The fronto-nasal sinuses are similar to those of N. paskoensis , but less expanded laterally (MDFE/LE: 0.61) ( Table 1) and less pneumatized. They are separated from the maxillaries and frontals, and from each other by a V-shaped notch. An extremely well-developed septum divides the nasal cavity into two chambers. The orbital notch has a distinct subelliptical outline, with the lower half directed distally. The sagittal crest is replaced by a medial concavity extending from the confluence of both nuchal crests to approximately the posterior third of the frontals. The zygomatic arches and descending processes of the maxillaries are much more developed than in other Neosclerocalyptus species, with the external margin of these structures with prominent ridges and rugosities extending to the ventralmost region of descending processes, at which point they are notably inclined towards the sagittal plane. The squamosal portions of zygomatic arches are subparallel to each other, differing from the condition of N. ornatus (Owen, 1845) and N. paskoensis , in which they diverge distally. The infraorbital foramina are larger than those of N. paskoensis and N. pseudornatus ( Ameghino, 1889) , and similar in size to those of N. ornatus , but more medially positioned. The occipital area is less expanded laterally than in N. paskoensis , but with greatly developed supraoccipital and paroccipital processes. The upper molariforms are almost undistinguishable from those of N. paskoensis . The mandible is robust, especially because of the greater anteroposterior width of the ascending rami, as in N. ornatus . The first lower molariform is simple and elongated longitudinally and the second molariform has incipient lobation, while the remaining molariforms are distinctly trilobed.
DESCRIPTION
Skull
The skull is similar in general morphology to that of N. paskoensis , with its convex dorsal profile, but it is more elongated anteroposteriorly ( Table 1); and less expanded laterally. There is less pneumatized of the fronto-nasal sinuses ( Fig. 1A, B, D View FIG ).
In lateral view ( Fig. 1C View FIG ), the parieto-occipital region is directed downwards as in N. paskoensis and E. tapinocephalus , although less markedly so than in these taxa. The zygomatic arches are much more robust than in the other species of Neosclerocalyptus (although proportionately shorter), with a rugose surface and a large tuberosity at its postero-inferior end. The zygomatic arches tend to be subparallel at the level of the squamosal portion, similar to the condition in Eosclerocalyptus proximus (Moreno & Mercerat, 1891) , and differing
from the condition in the other Neosclerocalyptus species in which these structures diverge distally. The orbital notches are clearly elliptical, with a notable dorsoventral development and the lower half markedly inclined distally. The descending process of maxillary is very robust and marked by a series of well-defined rugosities and ridges on its latero-external margin that have not been observed in any other Hoplophorinae Hoplophorini . The fronto-nasal sinuses are morphologically similar to those of N. paskoensis , with free margins straight, and differing from N. pseudornatus and N. ornatus in which these structures are recurved forming “loops”. These sinuses are separated from the maxillaries and frontals by a V-shaped notch, which is less evident than in N. ornatus .
In dorsal view, it was not possible to separate the cephalic shield from the skull. The parietals and frontals have a greater transverse diameter than in Ensenadan species. The sagittal crest present in N. pseudornatus and N. ornatus is replaced in this new species by a medial concavity extending approximately to the posterior third of the frontals, it then bifurcates to contact the origins of postorbital apophyses of frontals. Anteriorly, there is a notch separating the nasals from the rest of the skull, but it is not as deep as in N. ornatus .
In ventral view ( Fig. 1D View FIG ), the infraorbital foramina are remarkably large, and located more medially than in N. paskoensis and N. ornatus ; so they are more similar to those of N. ornatus and clearly different from those of E. tapinocephalus , N. pseudornatus and N. paskoensis . In these latter species, the infraorbital foramina are smaller and with a thicker lower margin. These foramina are located on a plane passing through M3, as in N. paskoensis . The lower third of the descending processes of the maxillary is inclined medially, more so than in N. ornatus and N. paskoensis . Both the upper and lower dental series tend to diverge anteriorly. The occipital area is not as expanded laterally as in N. paskoensis and N. pseudornatus , although it has a greater antero-posterior diameter. The palatal region anterior to M1 is not preserved,but the inferred morphology is very similar to that of the other species of Neosclerocalyptus . The M1 is simple, but slightly more elongated than in N. pseudornatus and N. ornatus . The M2 is already
trilobate, and the rest of molariforms (M3-M8) are
A new species of Neosclerocalyptus ( Xenarthra , Glyptodontidae ) from the Pleistocene of Argentina
not appreciably different from those of N. pseudornatus , N. ornatus and N. paskoensis .
In occipital view, the most prominent trait is the great development of supraoccipital and paraoccipital processes, which are the largest among the Hoplophorinae Hoplophorini .The foramen magnum has a more circular outline than in Ensenadan species, as in N. paskoensis . This character is probably related to the greater skull depth. As in N. ornatus , the choanae have a subrectangular outline and are taller dorsoventrally than the transverse width.
In frontal view ( Fig. 1A View FIG ), the nasal openings are large, and similar in outline to those of N. ornatus , but the free margins of nasals lack any curvature. The s eptum between both cavities is very well developed, more so than in N. ornatus and N. paskoensis , and homogeneously thick along nearly its entire dorso-ventral extension. The sinuses are morphologically similar to those of N. paskoensis , but less expanded transversally ( Table 1). Internally, the development of the trabecular tissue filling the sinuses is similar to that of N. ornatus , and not as dense as in N. paskoensis .
Mandible ( Fig. 1E, F View FIG ; Table 1)
The mandible is morphologically very similar to that of N. ornatus and N. pseudornatus , and more robust than that of N. paskoensis . The only remarkable characteristics are the greater lobation of M2 and the morphology of the coronoid apophysis, which is slightly higher than the condyle. In addition, the anterior margin of the ascending ramus has a series of ridges on its upper third, immediately below the coronoid apophysis. Lastly, sigmoid notch is slightly deeper than in N. ornatus , and more similar in this aspect to that of N. paskoensis .
Cephalic shield ( Fig. 1B View FIG )
As in other species of Neosclerocalyptus , the cephalic shield is rectangular in outline and formed by approximately 80 plates firmly sutured to each other. The central plates in the proximal half of the shield are the largest, with the rest decreasing in size laterally and posteriorly. Each plate bears a large circular or subcircular central figure with a smooth flat surface, although the largest ones
have a small central concavity. Each central figure is encircled by a row of figures, smaller but slightly more defined (especially those corresponding to the largest plates) than those of N. pseudornatus and N. ornatus . They are more rudimentary in shield area over the fronto-nasal sinuses, and almost disappear in ventralmost plates. The cephalic shield is not significantly different from other Neosclerocalyptus species in other aspects.
Humerus
The humerus is well preserved, showing the same basic morphology observed in the other species of Neosclerocalyptus (for more details see Zurita 2007).
GEOLOGICAL AND STRATIGRAPHICAL CONTEXT Ameghino (1889: 32-36) used the term Bonaerian as a synonym of “upper Pampean”. According to the views of this author ( Ameghino 1889: 32), the Bonaerian was characterized sedimentologically “by having in general lower proportion of lime and greater amount of sand, although at some points there are also large deposits of calcretes (‘toscas’);
however, it might be said to be less compact and
Zurita A. E. et al.
coherent than the lower pampean, and formed by a mass of pulverulent elements”. Another detailed mineralogical description of the “Bonaerian” was provided by Frenguelli (1957: 35): “Its grain is fine and homogeneous, its mass traversed by countless minute channels left by extremely fine roots, it is porous, non stratified, slightly calciferous, and its coloration is uniform light brown, generally with reddish hues.”
From a paleontological perspective, some authors (see Pascual et al. 1965, 1966; Marshall et al. 1984, among others) have not recognized the existence of the “Bonaerian”, arguing that its faunal composition was undistinguishable from that of the Lujanian s.s., and the use of this term was dropped as a result. Scillato-Yané (1979) and Reig (1981) were among the first authors to defend a differentiation within the Lujanian Mammal Age (sensu Pascual et al. 1965) and, following Ameghino, proposed its subdivision into two subages: Bonaerian and Lujanian. More recently, the Bonaerian Stage has been reincorporated into the chrono-biostratigraphic schemes, given that, although it is not clearly different from the Lujanian s.s (sensu Ameghino 1889) from a lithological perspective, it is clearly differentiable based on its biostratigraphy (see Cione & Tonni 1995, 1999; Tonni et al. 1999b). Currently, the Bonaerian is characterized biostratigraphically by the Megatherium americanum Cuvier, 1796 (Tardigrada Megatheriinae Owen, 1842) biozone (Cione & Tonni 1999, 2005).
Most of the geological units recognized for the Bonaerian are located within the modern limits of Buenos Aires province, and represented by the Buenos Aires Formation (see Parodi & Parodi 1952), and include the “Bonaerian Stage” and the “Belgranian Stage” (sensu Ameghino 1889) (Cione & Tonni 1999). Although with some uncertainty, the Bonaerian Stage could range chronologically from 0.40 to 0.13 Ma (Cione & Tonni 1999). MacFadden (2000) places the boundary between the Ensenadan and Bonaerian as slightly older, at approximately 0.6 Ma.
As we have already commented, the holotype of Neosclerocalyptus gouldi n. sp. (MCA 2010) consists of the complete skull, mandible, cephalic
shield and a right humerus of a specimen from Bonaerian sediments collected in the vicinity of San Andrés de Giles, Buenos Aires Province, and most likely corresponds to the Buenos Aires Formation. Paleomagnetic and chronostratigraphic studies performed near the Zárate locality indicate that the fossil bearing sediments could be correlatable with the Buenos Aires Formation (see Parodi & Parodi 1952), whose base is situated within the Matuyama Chron (reverse polarity), although most of it is included in the Brunhes Chron (normal polarity) ( Bobbio et al. 1986; Nabel 1993; Nabel et al. 1993, 2000).
This unit is sedimentologically very similar to the underlying formation (Ensenada Formation), but with less consolidated sediments ( Tonni et al. 1999b), and the predominance of smectites could be linked to greater water content at the time of sedimentation ( Nabel et al. 1993).
PALEOENVIRONMENTAL AND PALEOCLIMATIC CONTEXT
Sedimentological analyses of the Buenos Aires Formation show that, like the Ensenada Formation, is composed of alternating loess (deposited under cold arid conditions) and paleosoils (indicators of warmer and more humid climatic-environmental conditions), but with sediments less consolidated than in the underlying unit (Ensenada Formation) ( Orgeira et al. 1998; Tonni et al. 1999b; Nabel et al. 2000). In this sense, the Bonaerian seems to represent a predominantly cold arid or semiarid phase ( Cione & Tonni 2001), with brief interspersed warmer and more humid periods ( Tonni & Cione 1994). This interpretation is supported by both sedimentological and paleofaunal evidence; the predominant loess-type sedimentation of the uppermost Ensenada Formation indicates a shift toward colder and more arid conditions ( Tonni et al. 1999b; Nabel et al. 2000), also suggested by the presence of taxa clearly adapted to this type of environments (e.g., Lestodelphis halli (Thomas, 1921) , Microcavia Gervais & Ameghino, 1889 , Ctenomys Blainville, 1826 and Zaedyus Ameghino, 1889 ; Nabel et al. 2000); thus, an important faunal turnover occurred between the Ensenadan and the Bonaerian ( Ameghino 1889; Tonni et al. 1999b;
Cione & Tonni 2001).
A new species of Neosclerocalyptus ( Xenarthra , Glyptodontidae ) from the Pleistocene of Argentina
In this context, Fidalgo &Tonni (1983) described a micromammal association referable to the early-late Pleistocene from Punta Hermengo, Buenos Aires Province. These authors concluded that the entire sequence was deposited under predominantly cold arid conditions, and this hypothesis is supported by the presence of Zaedyus cf. Z. pichiy (Desmarest, 1806) , Tolypeutes cf. T. matacus (Desmarest, 1804) and Lagostomus Brookes, 1828 , among other taxa ( Tonni & Fidalgo 1982). However, more humid conditions, represented by limolitic sediments and lenses of calcrete clasts, have also been detected. Pardiñas & Lezcano (1995), in their analysis of a faunal association from northeastern Buenos Aires Province assigned to the Bonaerian, found an interesting group of taxa with warm humid ecological requirements (e.g. Lundomys Voss & Carleton, 1993 and Oxymycterus Waterhouse, 1837 ) along with others clearly adapted to dry or arid environments (e.g., Tolypeutes Illiger, 1811 and Microcavia ). Assuming that this entire fauna is synchronic, this co-occurrence could indicate the existence of mosaic-like environments, resulting from alternating dry cold phases and brief warm humid pulses. The presence of the latter is also supported by the presence of a chernozem-like paleosoil within the Pascua Formation ( Tonni & Fidalgo 1978).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |