
Parartemia veronicae
|
publication ID |
https://doi.org/ 10.5281/zenodo.199709 |
|
DOI |
https://doi.org/10.5281/zenodo.6200950 |
|
persistent identifier |
https://treatment.plazi.org/id/5D556922-6458-6545-FF0B-A0B8FDFBFC77 |
|
treatment provided by |
Plazi |
|
scientific name |
Parartemia veronicae |
| status |
|
Parartemia veronicae View in CoL
(Figs 4,7,12)
Parartemia View in CoL sp. d Timms and Savage, 2004, p25, 25, 32, 33, Campagna, 2007 Parartemia View in CoL sp. a. Chaplin, 1998
Parartemia View in CoL species B Clegg and Campagna 2006, p120.
Etymology. The species name honours Veronica Campagna who studied this species in Lake Yindarlgooda for her doctorate and who collected and/or raised many specimens of this species and of P. laticaudata sp. nov. for this study.
Holotype. Male, 110 km WSW of Norseman, Lake Johnson South, (32o 25’ 52”S, 120o 38’ 37”E), 20 January 2007, BVT, WAM 45242.
Allotype. Female, same collecting data as holotype, WAM 45243.
Paratypes. Three males, two females, same collecting data as holotype, WAM 45244; Three males, two females, same collecting data as holotype, AM P82977.
Other material. 10 males, 10 females, 137 km east of Norseman, unnamed salt lake east of Newman Rocks, (32o 07’ 01”S, 123o 11’ 15”E), 18 January 2007, BVT, WAM 45246; 5 males, 5 females, 39 km east of Norseman, unnamed roadside lake, (32o 04’ 10”S, 122o 08’ 14”E), 21 July 2003, BVT, WAM 45247.
Description. Male. Length 12.5 mm (head plus thorax 5.5 mm, abdomen 7 mm)
First antenna ( Fig. 12 View FIGURE 12 A) filiform, almost as long as basal antennomere of second antenna.
Second antenna. The two basal antennomeres ( Fig. 12 View FIGURE 12 A) fused at an angle of about 50o to body axis. Ventral margin with paired linear ventral processes, length about three times average depth. These processes with a rounded lateroventral corner not protruding laterally and with a prominent triangular medioventral corner; margin between the corners concave and medial margin convex and all margins lacking even minute spines. Medial process ( Fig. 12 View FIGURE 12 A) between ventral processes triangular and filling about the basal third of the space. Anterior processes ( Fig. 12 View FIGURE 12 A) triangular, length about three times average width and subequal to depth of medial margin sulcus of ventral processes. Distal antennomere ( Fig. 12 View FIGURE 12 A) of second antenna thin, cylindrical and tapering and about twice as long as basal antennomere.
Thoracic segments increasing in width 1st to 10th/11th. Thoracopods of Parartemia type but with a fewer posterior setae on endites 1+2, 3, exopodite and endopodite. 11th thoracopod without an epipodite, and first and last two smaller than remainder. Genital segments narrower than posterior thoracic segments and with 2nd genital segment oval in cross section, so giving a wider appearance than 1st abdominal segment from a dorsal perspective, but laterally a lower diameter than 1st abdominal segment.
Gonopods ( Fig. 12 View FIGURE 12 B) paired and about twice the diameter of tubular free apical part. Sharp triangular spine at base of free section and a small digitiform process subapically.
Abdominal segments increase in length and decrease further in diameter 1 to 6. 6th abdominal segment a little less than twice length of 1st segment.
Cercopods clothed with setae laterally and medially and subequal in length to 5th abdominal segment.
Female. Length 8.9 mm (head plus thorax 4.8 mm, abdomen 4.1 mm).
Head ( Fig. 12 View FIGURE 12 C) with first antenna filiform, shorter than eye plus peduncle. Second antenna about 1.5 times as long as eye plus peduncle, sausage shaped narrowing asymmetrically to an elongated apex. Naupliar eye prominent midway between the compound eyes. Labrum with a recurved spine.
Thoracomeres ( Fig. 12 View FIGURE 12 D,E) 1 to 9 normal, thoracomere 10 with large dorsolateral swellings and segment 11 with two smaller dorsolateral swellings, each somewhat sclerotised. Swellings of thoracomere 10 with a weak ridge horizontally near the segment margin. Brood pouch lateral lobes joined medioventrally and directed posteriorly at the gonopore, which is borne on a short tubular structure.
Thoracopods. Fifth thoracopod similar to that of male. First, second, ninth and tenth thoracopods smaller than third to ninth thoracopods. No eleventh thoracopod. First thoracopod (Fig. 7M) with all components, but with fewer posterior setae on endites 1+2, 3, endopodite and exopodite. Basal anterior setae of endites 4 and 5 longer than adjacent posterior setae and basal region of anterior setae of endite 1 about three times thicker than adjacent posterior setae and about as long. Tenth thoracopod (Fig. 7N) without an epipodite and praeepipodite, and a reduced exopodite protruding only a little more than the endopodite. Anterior setae all smaller than posterior setae. Posterior setae markedly reduced in numbers on endites 1+2 and 3 at about 8 and 3 respectively, though numbers on endopodite and exopodite less reduced at about 16 each.
Abdomen as in male, but covered in coarse denticles.
Variability. In males the relative length of the median process varies from about one quarter to almost one half of the depth of the medial margin of the ventral process, while the length of the anterior process also varies a little, though it is always a sturdy cone about twice as long as basal diameter. The ventral processes show a little variation in the degree of convexity of their ventral margins and in the prominence of the medioventral corner.
In subadult females of both collections east of Norseman, the thoracic swellings are not fully developed, and of course the brood chambers may be undeveloped. The lateral ridge on the lateral swellings of thoracomere 10 may be hard to detect, and the sclerotization of thoracopod 11 swellings may be incomplete. In general, the best developed of the swellings is the upper pair on thoracomere 11 and the least developed is the lateral swelling on thoracomere 10. In some specimens there is a weak swelling mid-dorsum on thoracomere 9.
Differential diagnosis. Both males and females of Parartemia veronicae sp. nov. are unspecialised without many of the special features seen in most other species. While it is usual for the female head to be undistinctive, the male head too has few remarkable features; the thorax lacks lateral lobes in both sexes and most female thoracomeres are little modified in other ways; females lack the 11th thoracopod as in most species, and other thoracopods are unremarkable; and the abdomen in both sexes is almost standard for Parartemia .
However the male is distinctive by having a triangular medial process, a feature shared only by P. acidiphila . In the latter species it is subequal in length to the depth of the ventral process and is notched at the apex, while in Parartemia veronicae sp. nov., the medial process is much shorter (<50%) than the ventral processes and has a single unnotched apex. The ventral processes are shaped differently from that in most species, what with the convex ventral margin, unfolded medial margin and especially the triangular medioventral corner. The later is reminiscent of that in P. triquetra , but in Parartemia veronicae sp. nov. is less prominent and lacks a spinous margin. These two species have a similar shaped frontal process, but can be further differentiated by the relative length of the first antenna― it is almost as long as the basal second antennomere in P. veronicae sp. nov., but only half this comparative length in P. triquetra . The frontal processes, while not unique, are certainly not massive and protruding beyond the ventral processes as in P. bicorna sp. nov., or hardly present as in Parartemia laticaudata sp. nov., or digitiform as in P. serventyi , P. contracta , P. longicaudata , and Parartemia boomeranga sp. nov., but they could be confused with those of P. triquetra , P. zietziana , P. informis and P. acidiphila .
The most distinctive feature in female Parartemia veronicae sp. nov. is the large tumidity on each side of thoracomere 10 and two small tumidities on each lateral surface of segment 11. Other species with thoracic tumidities include P. longicaudata , P. minuta , P. serventyi , P. yarleensis , P. purpurea sp. nov., P. boomeranga sp. nov., and P. laticaudata sp. nov.. The tumidities on these species are not on thoracomeres 10 and 11, nor in the combination (2 big; 4 small, all lateral) seen in Parartemia veronicae sp. nov. Having the brood chamber partly divided into lateral lobes is shared by a few species, including P. auriciforma , P. triquetra , P. yarleensis , P. purpurea sp. nov. and P. laticaudata sp. nov. The first three are separated by the presence of lateral thoracic lobes, P. purpurea sp. nov. has lateral flaps on thoracomeres 9 and 10, and P. laticaudata sp. nov. has three tumidities on thoracomere 9, one medial and two lateral (cf P. veronicae sp. nov., two tumidities on thoracomere 10, both lateral).
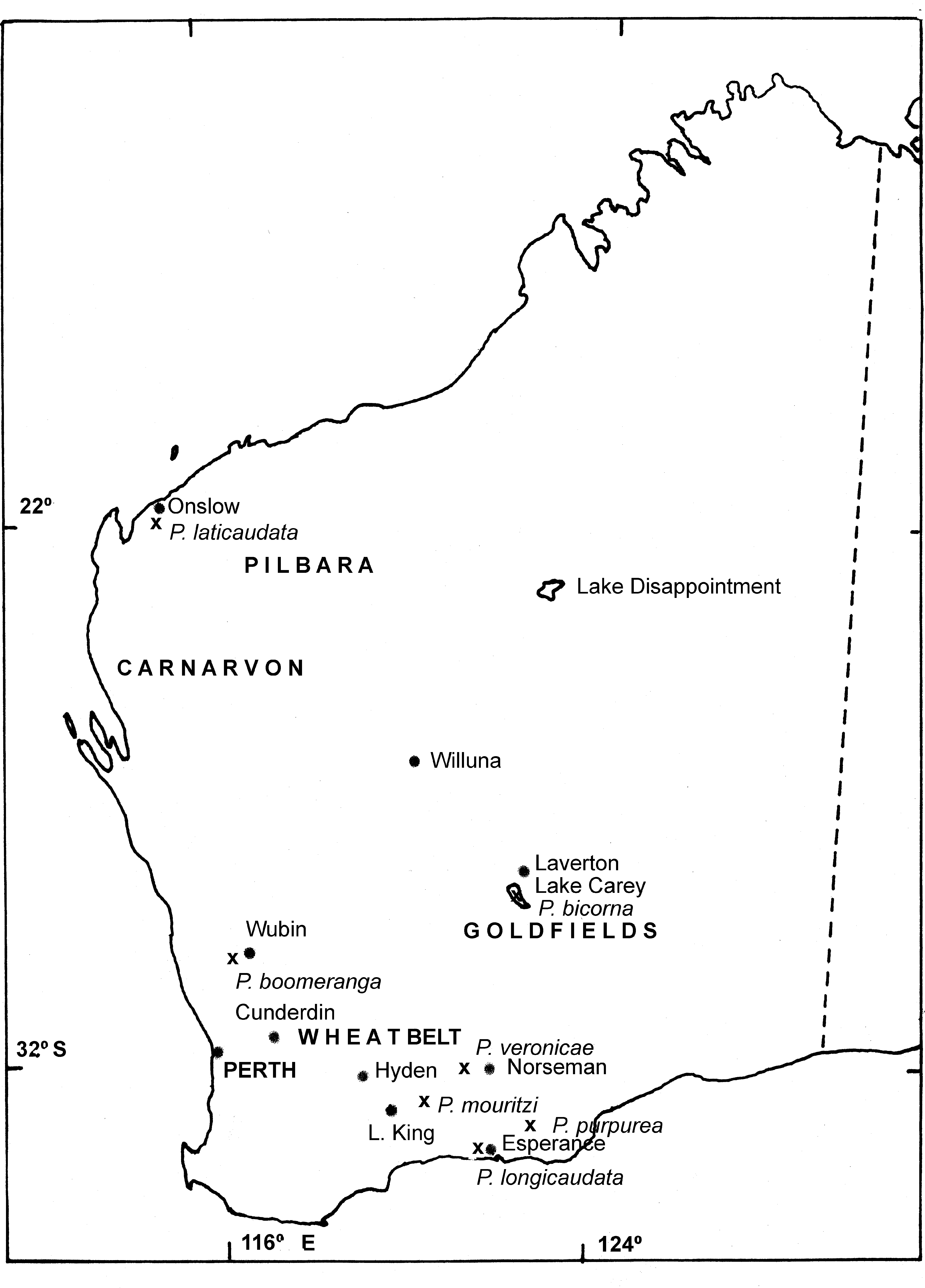
Distribution and ecology. Parartemia veronicae sp. nov. is known from the Goldfields from Willuna in the north to both and east and west of Norseman in the south (Timms et al. 2009)( Fig. 4 View FIGURE 4 ). Its known salinity range is 74 – 225 g /L ( Chaplin, 1998). Information on its ecology is given in Campagna (2007) (as Parartemia n sp. d) and Chaplin (1998) (as Parartemia sp. a). This species also has stress proteins and trehalose in its eggs to help survive in severe saline conditions (see comments on P. laticaudata sp. nov. above) ( Clegg and Campagna, 2006)

| WAM |
Western Australian Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.