
Tephritis vespertina (Loew, 1844)
|
publication ID |
https://doi.org/ 10.1080/00222933.2015.1010314 |
|
DOI |
https://doi.org/10.5281/zenodo.4330126 |
|
persistent identifier |
https://treatment.plazi.org/id/03F4879D-FF97-504F-9396-B621FC565C9A |
|
treatment provided by |
Carolina |
|
scientific name |
Tephritis vespertina |
| status |
|
Phytophagy: Tephritis vespertina View in CoL ; Chaetostomella cylindrica ; Acidia cognata (Tephritidae) ; Botanophila seneciella and Pegomya solennis (Anthomyiidae) .
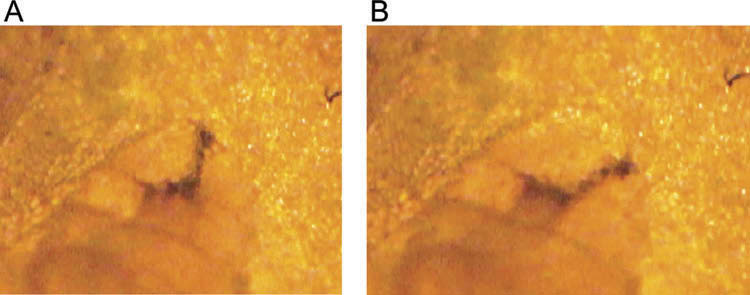
The larvae of all five species are found within living plant structures ( Table 1). T. vespertina , C. cylindrica and B. seneciella develop in different species of Asteraceae flowerheads, squeezing between florets and removing tissue ( Figures 38A and 38B View Figures 38 ). Access is similar in flowerheads and restricted++. Unlike the other two species, the larva of C. cylindrica completes development by tunnelling through the hard receptacle at the base of the flowerhead. A. cognata and P. solennis mine leaves of Tussilago farfara L. ( Asteraceae ) and Rumex acetosa L. ( Polygonaceae ), respectively, and feed on live mesophyll ( Figures 39 View Figure 39 and 40 View Figure 40 ). Access is restricted+++; however, access in leaves of R. acetosa is even more restricted as they are less than half as thick as those of T. farfara (<0.1–0.2 mm versus 0.4–0.6 mm. respectively, n = 3 leaves per plant species and measurements made at two equidistant points across the middle of each leaf).
All phytophages have mandibles with short, wide mandibular hooks; accessory teeth are usually present (little developed in B. seneciella ( Figure 41D–F View Figures 41 ), absent in C. cylindrica ) and elevator and depressor apodemes are more or less aligned one under the other, with the result that the base is crescent-shaped in lateral view ( Figure 41A–G View Figures 41 ). Cibarial ridges are absent and dental sclerites reduced or absent. The intermediate sclerite is completely sclerotised and, in the tephritids, abuts smoothly with a forward projection of the basal sclerite ( Figure 41A–C View Figures 41 ). The labial sclerite is developed and, in lateral view, appears as a curved, ventral bar between the mandible and the intermediate sclerite ( Figure 41C and 41E View Figures 41 ). The valve at the end of the ventral cornu is weak (i.e. not sclerotized) and windows and areas of poor sclerotisation are present in both the dorsal and ventral cornua ( Figure 41A–D and 41G View Figures 41 ). The pseudocephalon is dome-shaped ( Figure 42 View Figure 42 ), with a slightly elongate or oval-shaped oral cavity, cirri are absent and the ventral margin has a transluscent, curved oral plate, sometimes with a small, sclerotised section (entirely transluscent in P. solennis and C. cylindrica ) and slightly shorter than the length of the head skeleton.
Sweeping is less pronounced and consists of downward rather than sideways movement of the middle body compartment. As one lunge ends:
● abdominal segments 8–2 are stationary;
● abdominal segment 1, the metathorax, mesothorax and the prothorax to the level of the anterior spiracles are extended;
● the prothorax beyond the spiracles is inclined;
● the pseudocephalon is extended dorsally and collapsed and/or folded into the prothorax ventrally and the head skeleton is inclined and the mandibles depressed;
A new lunge begins when:
● the head skeleton pivots up by about 10–20 degrees with respect to the horizontal position ( Table 2) and the mandibles rise; dorsally, the pseudocephalon folds slightly and ventrally it unfolds ( Figure 43A View Figures 43 );
● as pivoting ends and the mandibles are against plant tissue, the head skeleton starts pivoting in the opposite direction which pulls the mandibles down across plant tissue; the mandibles pivot down by up 60 degrees ( Table 2), and dorsally, the pseudocephalon unfolds and ventrally folds ( Figure 43B View Figures 43 );
● for most of the downward pivot, the mandibles do not depress until the end of the pivot when they depress to their limit; rasped material fills the oral cavity and is sucked in by the pump in the head skeleton.
Flowerhead feeders: body short and broad ( Figures 38 View Figures 38 , 44–46 View Figure 44 View Figure 45 View Figure 46 , Table 3). Spicules are extensive, except for being reduced on the middle abdominal segments, metathorax and mesothorax and only present on antero-dorsal and antero-ventral margins of the prothorax. T. vespertina with mandibular muscle apodemes straight, not curved ( Figures 41A View Figures 41 ) and mandible almost as long as wide with one accessory tooth. Mandible are longer than wide in B. seneciella and C. cylindrica ; B. seneciella has a tapered apex to the mandibular hook ( Figure 41 View Figures 41 (f)). In living larvae, the upper margin of the dorsal cornu is bent inwards along the window and the front body compartment is able to rotate 360 degrees. In all three species, holes are made in floret tissues, the margins of which decay and the larva can be surrounded by moist to wet decayed material on which it may feed ( Figure 38A View Figures 38 ). Larvae start development by feeding towards the base of florets, but readily reverse direction. Lunge times in T. vespertina were 0.4–0.7 sec per lunge and 0.5–0.6 sec per lunge in B. seneciella ; lunge times were not measured in C. cylindrica ( Table 2).
Leaf miners: body elongate ( Figures 38 View Figures 38 , 47 View Figure 47 and 48 View Figure 48 , Table 3). Spicules are extensive and circumvent segment borders, especially on the anterior abdominal segments and the metathorax. Unlike A. cognata which is symmetrical, in P. solennis the pseudocephalon is asymmetrical with a longer lower side, i.e. when feeding the shorter margin faces the upper epidermis of the leaf ( Figure 49 View Figure 49 ). The pseudocephalon in P. solennis is more tapered than in A. cognata , such that the antennae are approximated whereas, in A. cognata , antennae are separated by about 1.5× their width. The lateral margins of the pseudocephalon in P. solennis have three square-shaped studs on the long side and six on the short side ( Figure 49 View Figure 49 ); studs are absent in A. cognata . The mandibular muscle apodemes curve backwards in both species. The mandibles are symmetrical in A. cognata with one accessory tooth each, but asymmetrical in P. solennis with the longer, lower mandible having a strongly curved, apical mandibular hook and two, short, accessory teeth and the lower mandible with a similar strong, curved apex to the mandibular hook and three, elongate accessory teeth ( Figure 50 View Figure 50 ). The prothorax in P. solennis has a conspicuous, lozenge-shaped, mid-ventral projecting bar ( Figure 42 View Figure 42 ). These larvae excavate leaf tissue in very different ways.
Acidia cognata : feeding on its side this larva removes tissue in a series of long, crescent-shaped feeding tracks. Each track consists of succeeding numbers of arcshaped excavations of tissue; both tracks and arcs are traceable by remnants of tissue left on the upper epidermis ( Figure 40 View Figure 40 ). These traces are not frass, which is otherwise scattered through the mine as lumps of moist, brown material ( Figure 39 View Figure 39 ). Track direction is influenced by barriers, such as leaf veins, leaf edges, damaged areas and presumably, other larvae in the leaf. Arcs are made by the front compartment sweeping down across the mine face during which lunging occurs continuously with each lunge removing a line of tissue. To lunge, the rear compartment grips the upper and lower layers of the leaf, the middle compartment extends and the front compartment lifts and the mandibles elevate slightly and are placed against the mine face. The head skeleton pivots down and the mandibles depress slightly and a line of tissue is removed (Film 17). The prothorax moves slowly down across the mine face with the head skeleton pivoting continuously; a small arc so filmed, took about 74 seconds to create and involved about 114 head skeleton pivots, i.e. lunge time was about one lunge per 0.6 sec. At the end of an arc, the larva moves into the excavated space, lifts the prothorax and begins to make another arc and so, a track gradually forms.
Pegomya solennis : the larva also feeds on its side but instead of tracks and arcs, oval-shaped cavities are formed in the leaf, but little tissue is left behind to indicate the pattern of feeding. To start lunging, the larva lifts the front compartment and the mandibles elevate slightly and are pressed against the mine face (Film 18, Figure 43A View Figures 43 ). The head skeleton pivots and a line of tissue is removed by the mandibles which depress slightly in the process. At the limit of depression, the fleshy, prothoracic bar is adjacent to the mandibles ( Figure 43B View Figures 43 ). The front compartment then lifts and the process repeats. Lunge times, the time for a line of tissue to be removed, were about twice as fast as A. cognata and varied from about 0.1 to 0.3 second. When a space has been created in front of the larva, the prothorax extends, the head skeleton pivots and protracts, and the mandibles elevate and grip the mine face. A peristaltic wave pushes the larva up against the mine face and another cycle of lunges start (Film 18). Blotch-shaped excavation occurs via the direction of lunging altering slightly. This depends where the mandibles grip the mine face prior to peristalsis; if below the longitudinal axis of the body, then lunging is in that direction, if above, then in that direction.










No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |