
Phragmatopoma digitata Rioja, 1963
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4845.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:D737FF5C-63F8-4E80-A605-789A5FEB1B2C |
|
DOI |
https://doi.org/10.5281/zenodo.4407388 |
|
persistent identifier |
https://treatment.plazi.org/id/977F915B-FFB6-FFD7-FF7C-FF3EFB55FE3F |
|
treatment provided by |
Plazi |
|
scientific name |
Phragmatopoma digitata Rioja, 1963 |
| status |
|
Phragmatopoma digitata Rioja, 1963 View in CoL
Figures 5 View FIGURE 5 A–L, 12B
Phragmatopoma moerchi digitata Rioja, 1963: 200–201 View in CoL , figs. 106–112. Type locality: La Ventosa Beach, Salina Cruz, Oaxaca, Mexico.
Phragmatopoma digitata View in CoL .— Achari 1974: 451 (world comparison of sabellariids); Salazar-Vallejo & Londoño-Mesa 2004: 51 (checklist of polychaetes from tropical eastern Pacific).
Phragmatopoma virgini View in CoL (not Kinberg, 1867).— Kirtley 1994: 37–40, figs. 2.4.7 (La Ventosa, Salina Cruz, Oaxaca).
Phragmatopoma View in CoL sp. 1 Chávez-López & Cruz-Gómez 2019: 161 ( Agua Blanca , Oaxaca, Mexico, 0.2 m, on rocks and sand) .
Type material. Neotype: ECOSUR0227 View Materials , La Ventosa , Salina Cruz, 16°10’46.1”N, 96°09’25.4”W, Oaxaca, Mexico, southern Mexican Pacific; 0.2–0.5 m, May 21, 2000, coll. J. R. Bastida-Zavala & S.I. Salazar-Vallejo; GoogleMaps Paraneotypes: ECOSUR0228 View Materials , 479 spec. from La Ventosa, Salina Cruz, same as neotype. GoogleMaps
Additional material: 99 specimens. Jalisco: ECOSUR-P3094, six spec. (Melaque, 19°12’59.6”N, 104°42”51.3”W, December 1, 2004). GoogleMaps Guerrero: ECOSUR-P3095, two spec. (La Condesa, Acapulco , 16°51’23.3”N, 99°52’08.3”W, November 27, 1999, coll. S.I. Salazar-Vallejo) GoogleMaps ; ECOSUR-P3096, one spec. (Los Cantiles, Acapulco , 16°51’35”N, 99°54’23”W, on bivalve shells, 5 m, May 26, 2000, coll. A. Medina-López) GoogleMaps ; UMAR-Poly-970, four spec. ( Punta Maldonado , 16°19’36.5”N, 98°34’15.6”W, November 8, 2017, coll. J. R. Bastida-Zavala & P. Hernández-Moreno) GoogleMaps . Oaxaca: UMAR-Poly-971, one spec. ( Chacahua Lagoon , 15°57’59”N, 97°40’44”W, intertidal, on rocks, April 13, 2007) GoogleMaps ; UMAR-Poly-972, one spec. ( Chacahua Lagoon , 15°57’59”N, 97°40’44”W, on algae, April 13, 2007) GoogleMaps ; UMAR-Poly-973, two spec. ( Puerto Ángel , 15°39’49”N, 96°29’40”N, on spring piles, May 20, 2007, coll. F. Cortés-Carrasco & S. Rodríguez-Hernández) ; UMAR-Poly-974, one spec. ( Estacahuite , 15°40’06”N, 96°28’52”W, on coral Pocillopora , coll. S.I. Salazar-Vallejo & J.A. de León-González, April 17, 2008); UMAR-Poly-975 GoogleMaps , 54 spec. ( Agua Blanca , 15°43’57”N, 96°48’42”W, on sand, September 9, 2010, coll. B. Moreno-Dávila) GoogleMaps ; UMAR-Poly-976, one spec. ( Corralero Lagoon , 16°12’54”N, 98°11’11”W, on rocks, April 18, 2013, coll. E. Pérez-Pacheco) GoogleMaps ; UMAR-Poly-977, five spec. ( Puerto Ángel , 15°39’49”N, 96°29’40”N, on coral rock, April 20, 2013, coll. E. Lazo-Zúñiga) ; UMAR-Poly-978, 13 spec. ( Chacahua Lagoon , 15°57’59”N, 97°40’44”W, on algae, April 13, 2016) GoogleMaps ; UMAR-Poly-979, two spec. ( Agua Blanca , 15°43’57”N, 96°48’42”W, 0.2 m, on rocks, March 17, 2017, coll. K.J. Humara-Gil & Y. Chávez-López) GoogleMaps .
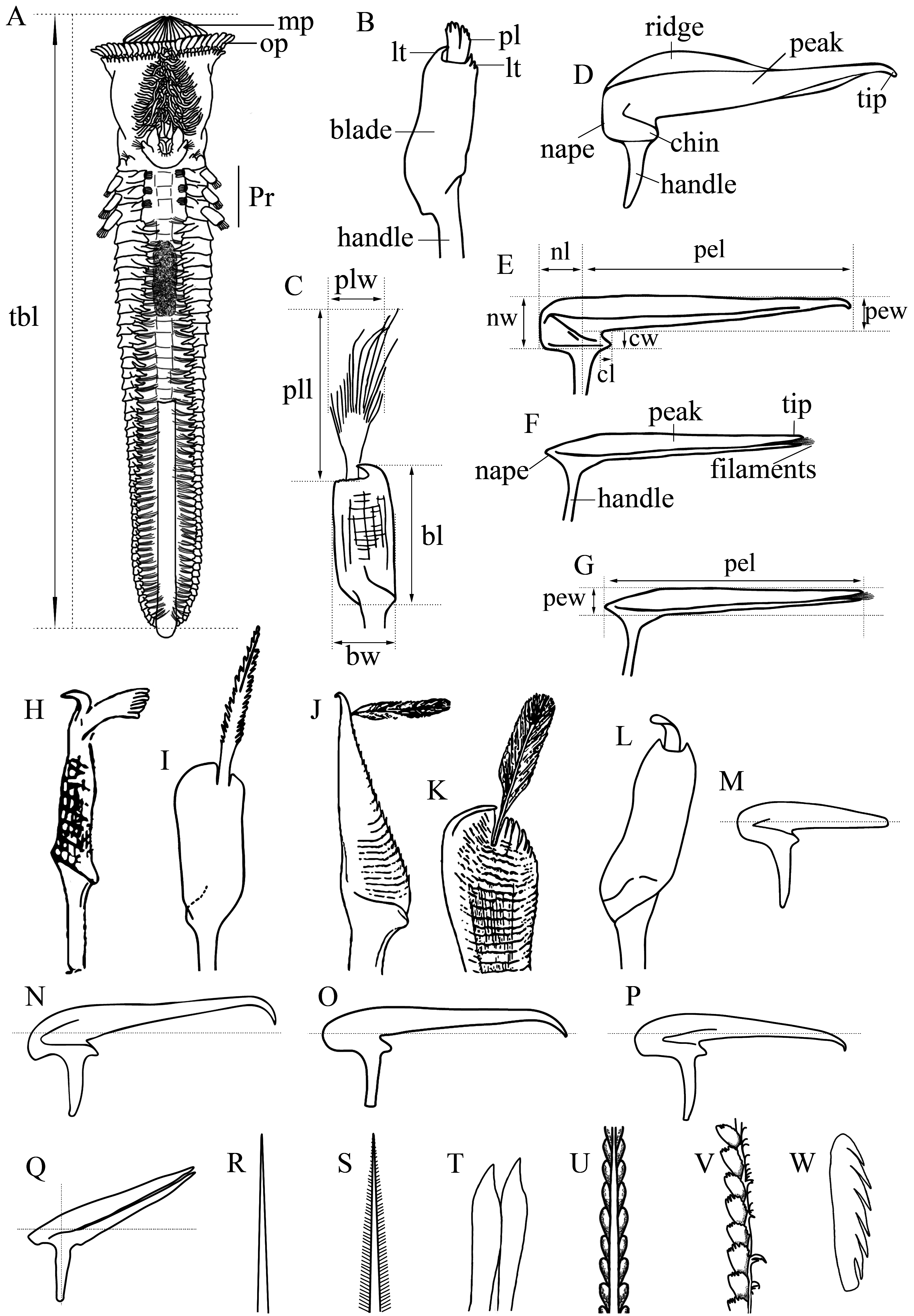
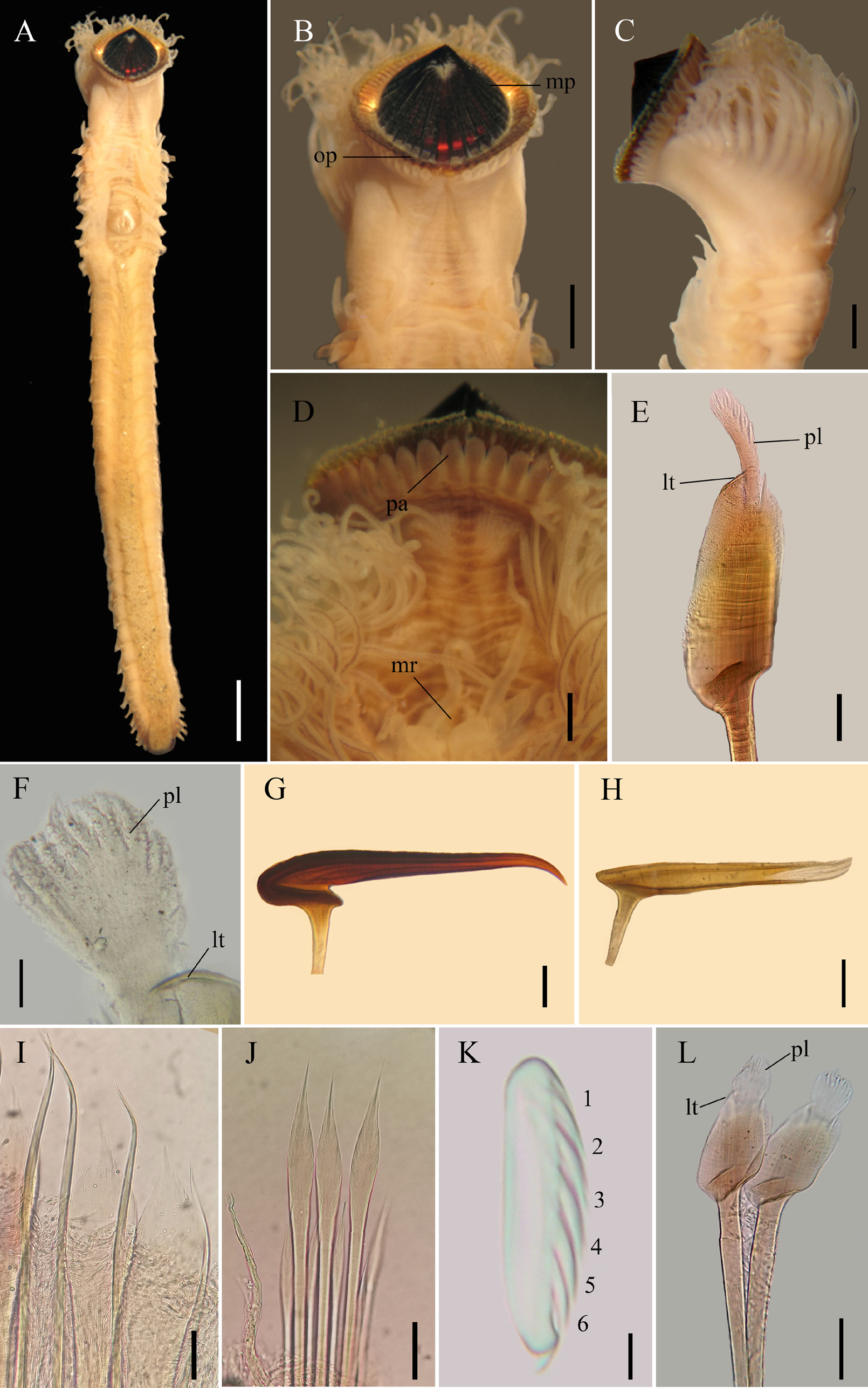
Description. Color pattern of preserved specimen s. Body pale yellow ( Fig. 5A View FIGURE 5 ). Outer paleae with blade and handle amber; median plume translucent ( Fig. 5E, F View FIGURE 5 ). Middle paleae cherry with the handle amber ( Fig. 5G View FIGURE 5 ). Inner paleae amber (Fig. H). Opercular papillae pale yellow with brown points in the center ( Fig. 5D View FIGURE 5 ). Median ridge with light brown eyespots ( Fig. 5D View FIGURE 5 ). Tentacles pale yellow with a light brown transversal line ( Fig. 5D View FIGURE 5 ). Building organ and branchiae pale yellow. Parathoracic chaetae translucent ( Fig. 5 View FIGURE 5 I–J). Abdominal neuropodia with two series of brown points. Abdominal neurochaetae and uncini translucent ( Fig. 5K View FIGURE 5 ). Caudal peduncle yellowish to translucent toward distal part, basal region with lateral brown points.
Body. Complete specimen of 17 mm total length; parathoracic region 3 mm wide; 32 abdominal segments; caudal peduncle 3.8 mm long ( Fig. 5A View FIGURE 5 ).
Operculum. Opercular crown and opercular stalk completely fused. Opercular crown conic and oval, slightly protruding in lateral view ( Fig. 5 View FIGURE 5 A–D). Three rows of paleae, only two visible: 70 outer paleae, 29 middle and inner paleae. Outer paleae geniculate with a pair of heterodont teeth, one straight, and the other slightly curved ( Fig. 5E View FIGURE 5 ); flat blade almost three times longer than wide, serrated margin and without transversal thecae; median plume short, ½ as long as blade, rounded, longer than wide, filamentous with thin filaments, ¼ as long as plume ( Fig. 5 View FIGURE 5 E–F). Middle paleae strongly geniculate with straight peak, rough surface with transversal thecae; sub-circular nape, straight, serrated surface, wider than peak, ¼ as long as peak; chin slightly longer than wide, margin serrated; sharp tip markedly falcate, smooth margin ( Fig. 5G View FIGURE 5 ). Inner paleae strongly geniculate with elevated serrated peak, eight times longer than wide; nape smooth; tip with filaments ( Fig. 5H View FIGURE 5 ). Papillae long, oval. Oral tentacles unbranched. Median ridge short, 1/5 as long as opercular stalk, with marginal eyespots ( Fig. 5D View FIGURE 5 ). Median organ absent. Building organ ‘U’- shaped.
Thorax. Chaetiger 1 with a pair of neuropodia, bipinnate chaetae. Chaetiger 2 with three pairs of conical cirrus, neuropodia with bipinnate chaetae and a pair of branchiae.
Parathorax. Three parathoracic segments ( Fig. 5A View FIGURE 5 ). Chaetigers with a pair of branchiae. Notopodia with lanceolate chaetae interspersed with long capillary chaetae ( Fig. 5I View FIGURE 5 ). Neuropodia with lanceolate chaetae interspersed with small lanceolate chaetae ( Fig. 5J View FIGURE 5 ); neurochaetae thinner than notochaetae.
Abdomen. Segments with a pair of branchiae decreasing in size towards posterior segments; absent in the last chaetigers. Neurochaetae verticillate of different length. Notopodia with a series of uncini with six pairs of teeth ( Fig. 5K View FIGURE 5 ).
Caudal region. Caudal peduncle elastic, smooth, without segmentation.
Tubes. Phragmatopoma digitata builds tubes with thick sand grains and some fragments of shells ( Fig. 12B View FIGURE 12 ).
Variation. Body measurements varied from 4.5–20 mm total length, parathoracic region 0.5–3 mm wide and caudal peduncle 0.8–3.8 mm long. The number of abdominal segments varied from 17–33. The body color varied with size body, founding translucent and yellow color, in small specimens (<2 mm total length) and pale yellow in specimens longer. Small specimens have the basic morphology of paleae but with lighter shades and translucent paleae ( Fig. 5L View FIGURE 5 ).
Habitat. Phragmatopoma digitata can build large aggregations on rocks, as in La Ventosa, Oaxaca (Salazar-Vallejo com. pers.), from the intertidal zone to 5 m. Also, this species may be found on pier pilings, as in Puerto Ángel, Oaxaca, and can be associated with the coral Pocillopora or with algae.
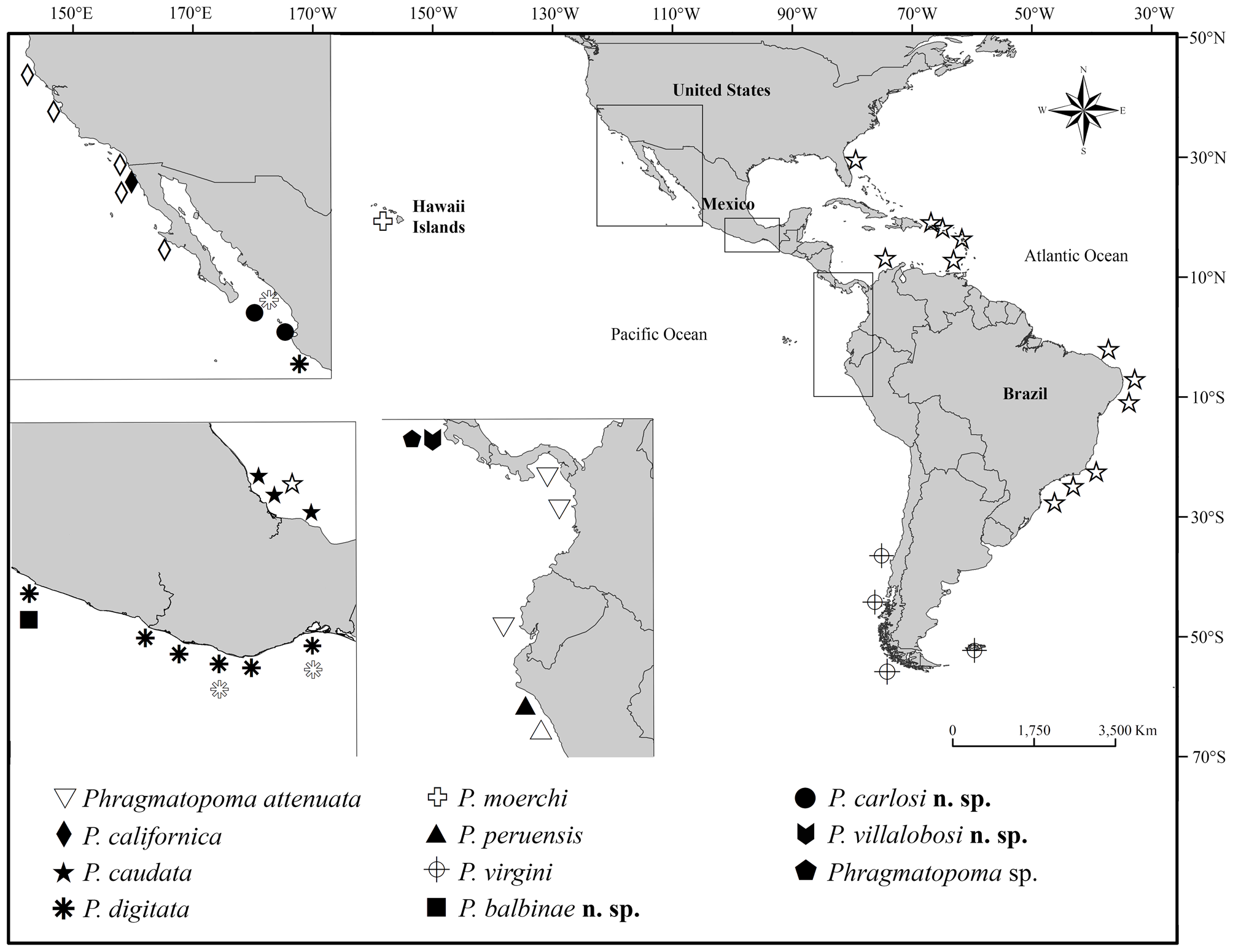
Distribution. Only known from Jalisco, Guerrero and Oaxaca, Mexico. According to Rioja (1942), it is also possibly distributed in Mazatlán, Sinaloa ( Fig. 13 View FIGURE 13 ).
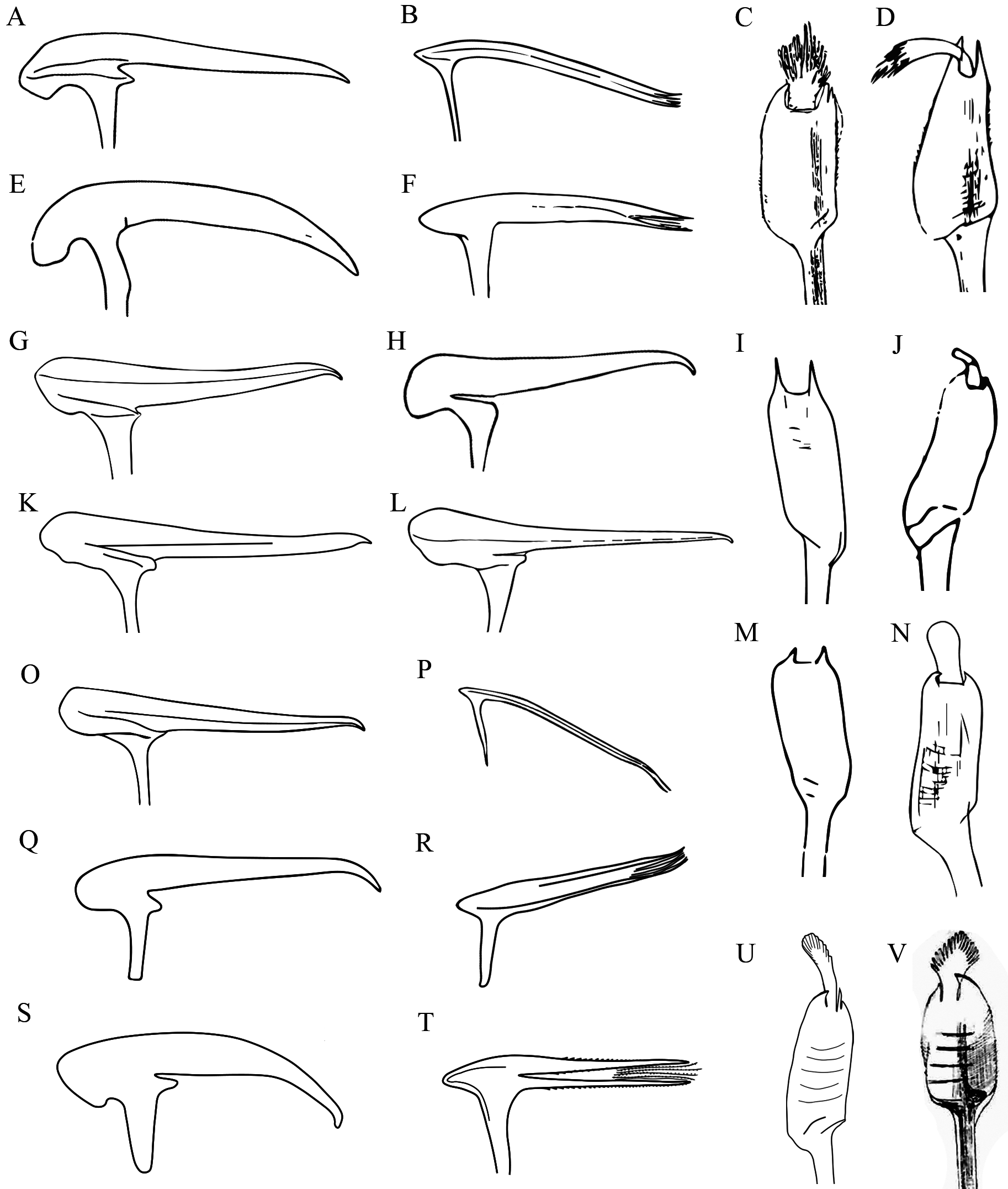
Remarks. Phragmatopoma digitata was described from La Ventosa, Salina Cruz, Oaxaca, Mexico ( Rioja 1963), with filamentous median plume in outer paleae ( Fig. 6 View FIGURE 6 C–D); middle paleae with decurrent nape, slightly declined and convex peak, without chin ( Fig. 6E View FIGURE 6 ); and inner paleae straight ( Fig. 6F View FIGURE 6 ). After its discovery, P. digitata has not been re-described or recorded; moreover, the type material is lost.
Kirtley (1994) synonymized P. digitata (La Ventosa, Salina Cruz), P. peruensis (Salaverry, Peru) and P. moerchi (Honolulu, Hawaii) with P. virgini (Straits of Magellan, Chile), expanding its distribution from cold waters of the Chilean Patagonia to tropical waters of southern Mexico, and including one locality of Baja California (northern Mexico).
The description of P. virgini is very confusing and vague. Kinberg (1867) described P. virgini with an elongate spine in outer paleae but did not include any illustration. Johansson (1926) illustrated the outer paleae of P. virgini without median plume, and Hartman (1944) revised the type material of this species, described and illustrated the outer and middle paleae. According to Hartman (1944), P. virgini have outer paleae with flat blade, almost three times longer than wide, without median plume or indications of presenting any membranous structure ( Fig. 6I View FIGURE 6 ); middle paleae with slightly concave peak, straight nape and curved tip ( Fig. 6G View FIGURE 6 ), and uncini with six pairs of teeth. Kirtley (1994) illustrated the outer paleae of the holotype of P. virgini with flat blade and oval median plume smooth ( Fig. 6J View FIGURE 6 ), and middle paleae with elevated peak, decurrent nape and falcate tip ( Fig. 6H View FIGURE 6 ). However, he also illustrated outer and middle paleae of topotype material with different morphology, presenting outer paleae without median plume ( Fig. 6M View FIGURE 6 ) or oval plume, and middle paleae with straight peak and nape, and curved tip ( Fig. 6K View FIGURE 6 ) as in Hartman’s (1944) description.
Phragmatopoma virgini , P. peruensis , P. moerchi , and P. digitata varied principally in morphology of median plume of outer paleae. Phragmatopoma moerchi has a rectangular lacerate plume ( Hartman 1944, Fig. 2B View FIGURE 2 ); P. peruensis has a smooth oval plume ( Hartman 1944, Fig. 6N View FIGURE 6 ); P. virgini may have a plume similar to that of P. peruensis ( Kirtley 1994, Fig. 6J View FIGURE 6 ) or the plume can be absent ( Hartman 1944, Fig. 6I View FIGURE 6 ; Kirtley 1994, Fig. 6M View FIGURE 6 ); and P. digitata is distinguished by sub-quadrangular filamentous plume ( Rioja 1963, Fig. 6 View FIGURE 6 C–D). Kirtley (1994) mentioned that outer paleae morphology, particularly that of median plume, could be changed by abrasion; however, the differences observed in the Oaxaca and Peru populations are constant in specimens, so these cannot be attributed to abrasion. In addition, the four species of the genus Phragmatopoma show variations in middle paleae such as: sub-circular nape in P. virgini ( Hartman 1944, Fig. 6G View FIGURE 6 ) P. moerchi ( Fig. 6O View FIGURE 6 ) and P. digitata ( Figs. 5G View FIGURE 5 , 6Q View FIGURE 6 ), and sub-quadrangular in P. virgini ( Kirtley 1994, Fig. 6H View FIGURE 6 ) and P. peruensis ( Fig. 7G View FIGURE 7 ); the straight peak in P. peruensis ( Fig. 6L View FIGURE 6 ) and P. digitata ( Fig. 6Q View FIGURE 6 ), declined in P. moerchi ( Hartman 1944, Fig. 6O View FIGURE 6 ), elevated in P. virgini ( Kirtley 1994, Fig. 6H View FIGURE 6 ); and the falcate tip in P. virgini ( Hartman 1944, Fig. 6G View FIGURE 6 ) and P. moerchi ( Fig. 6O View FIGURE 6 ), markedly falcate in P. digitata ( Fig. 6Q View FIGURE 6 ), and curved in P. peruensis ( Fig. 6L View FIGURE 6 ). Finally, the inner paleae are also differ among species, straight in P. peruensis ( Fig. 7H View FIGURE 7 ), elevated in P. digitata ( Fig. 6R View FIGURE 6 ), and decurrent in P. moerchi ( Fig. 6P View FIGURE 6 ) (Table I).
The examined specimens are similar to the ones described by Rioja (1963) in the morphology of inner and outer paleae, with slight differences in the size of filamentous in median plume, longer in Rioja’s (1963) specimens, ~1/3 of total length of median plume ( Fig. 6 View FIGURE 6 C–D), and short in my specimens, ~1/4–1/5 of total length of median plume ( Fig. 5 View FIGURE 5 E–F, 6U). However, the principal difference with the original description is in middle paleae. The Rioja’s (1963) specimens have a peak declinate, nape decurrent, tip rounded and lack a chin ( Fig. 6E View FIGURE 6 ), while my specimens have peak straight and not declined, nape straight and not decurrent, sharp tip and not rounded and have chin ( Fig. 5G View FIGURE 5 , 6Q View FIGURE 6 ).
Rioja (1963) did not include any information about body morphology of P. digitata , therefore, it is impossible to determine other similarities or differences between his specimens and mine, including topotypic specimens. The differences observed in the morphology of middle paleae, described by Rioja (1963), may have been influenced by different factors like: 1) resolution of optic device used; 2) some assembly error, influencing the visual perception of the shape of paleae; or 3) intraspecific variation.
Based on the morphological differences among Phragmatopoma species, the synonymies cannot be sustained and consequently, P. digitata must be regarded as a distinct species. Herein, the neotype of the species is established based on topotypical lots, corresponding to 82% (480 spec.) of total revised material of this taxon.


| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Phragmatopoma digitata Rioja, 1963
| Chávez-López, Yessica 2020 |
Phragmatopoma
| Chavez-Lopez, Y. & Cruz-Gomez, C. 2019: 161 |
Phragmatopoma virgini
| Kirtley, D. W. 1994: 37 |
Phragmatopoma digitata
| Salazar-Vallejo, S. I. & Londono-Mesa, M. H. 2004: 51 |
| Achari, G. P. K. 1974: 451 |
Phragmatopoma moerchi digitata
| Rioja, E. 1963: 201 |