
Simoclaenus sylvaticus Muizon & Cifelli, 2000
|
publication ID |
https://doi.org/ 10.5252/geodiversitas2019v41a25 |
|
publication LSID |
urn:lsid:zoobank.org:pub:4EF01B8A-BA1D-4F14-9432-CA8754D765D2 |
|
DOI |
https://doi.org/10.5281/zenodo.3704860 |
|
persistent identifier |
https://treatment.plazi.org/id/291A87B5-FFEB-1E15-FF75-F4791EEDFEF8 |
|
treatment provided by |
Valdenar |
|
scientific name |
Simoclaenus sylvaticus Muizon & Cifelli, 2000 |
| status |
|
Simoclaenus sylvaticus Muizon & Cifelli, 2000
EMENDED DIAGNOSIS. — Dental formula: I?/?, C 1/1, P 4/4, M 3/3; relatively large short-snouted kollpaniine characterized by the strong mesiodistal compression of its lower molars and premolars, which is correlatively observed on the M2; upper canine transversely compressed; P1 vertically implanted; P2 with a small protoconal bulge on lingual side of paracone; P3-4 with well-developed protocone (slightly larger on P4); M1 much smaller than M2; M2 almost square in occlusal outline but distinctly asymmetrical as in Molinodus ; p4 with a well-developed metaconid medial to protoconid; molars with a large metaconid, posterior to the protoconid and with a posterior projection that partially fills the talonid basin; protoconid distinctly smaller than in Andinodus; posterior wall of trigonid sigmoid in occlusal view; paraconid and paracristid smaller than in Andinodus; large hypoconid with a labial slope less vertical than in Andinodus; cristid obliqua meets trigonid on lingual edge of protoconid.
Ascending process of the maxilla short, high, and erected, premaxilla-maxilla suture subvertical in lateral view; frontal-maxilla suture present (therefore absence of nasal-lacrimal suture) anterior opening of infraorbital canal at level of P3. Rostrum, blunt and short as indicated by cheek teeth mesiodistal compression, implantation of anterior premolars, and subvertical premaxilla-maxilla suture.
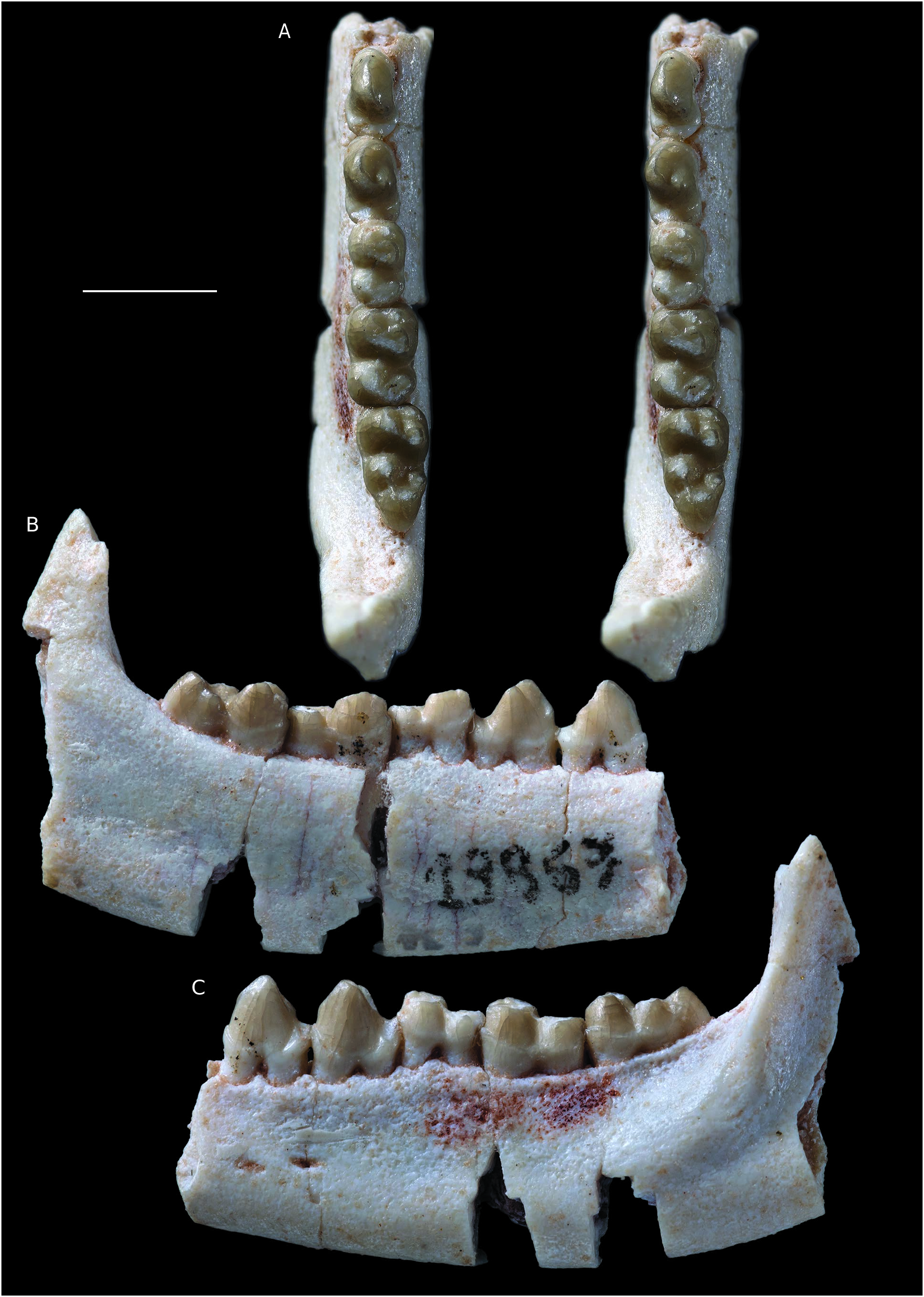
HYPODIGM. — The type specimen, MHNC 8332, a fairly complete right dentary with the alveoli of c, p1, the roots of p2, the alveolus of p3, and p4-m3. The dentary is lacking the anterior region with the incisor alveoli and most of the coronoid process; MHNC 8348 View Materials , a right maxilla with M1 lacking the parastylar region and part of the paracone and M2-3 complete ; MHNC 13868 View Materials , a right maxilla with root of the canine and P1-P4 ; MNHC 13872 , a partial mandible with alveolus of p1, roots of p2-3and p4-m1 ; MHNC 13876 View Materials a right maxilla fragment with M1-M2 .
DESCRIPTION
Three new specimens are referred to this species. MHNC 13872 is a mandible fragment with the alveolus of p1, the roots of p2-3 and p4-m1 ( Fig. 7 View FIG ). The alveolus of p1 is vertical as in the holotype of Simoclaenus sylvaticus (MHNC 8332) ( Muizon & Cifelli 2000), indicating that the tooth was not procumbent in contrast to the condition observed in Molinodus (see above). This feature, shared by the holotype of Simoclaenus sylvaticus and the new specimen clearly supports the referral of the latter rather than to Molinodus suarezi , the p1 of which is procumbent ( Fig. 1A, B View FIG ). However, the major characteristic of this specimen lies in its p4, which is markedly compressed mesiodistally as is observed on the holotype. As in the latter, the metaconid is placed lingually to the protoconid and is not shifted posteriorly as is observed in Molinodus suarezi ( Fig. 1D View FIG ). Furthermore, the p4 of MHNC 13872 has the same strongly bulbous morphology as that of the holotype. The m1 is relatively worn but does not depart significantly from that of the holotype in terms of morphology.
However, the two specimens differ in their size and the new specimen described here is distinctly smaller than the holotype ( Table 2). As a matter of fact, the p4 and m1 of the new specimen are in average 10% smaller than those of the holotype (mean of the four ratios of Table 2). Although significant, this difference could be interpreted as related to individual size variation. The other large “condylarth” to which this specimen could be related is Molinodus suarezi , although this taxon is clearly smaller than Simoclaenus sylvaticus . However, the p4 observed on the mandible of M. suarezi described above ( Fig. 2A View FIG ) is so different from that of MHNC 13872 that we rather favor a strong individual size variation in S. sylvaticus than morphological variation in M. suarezi .
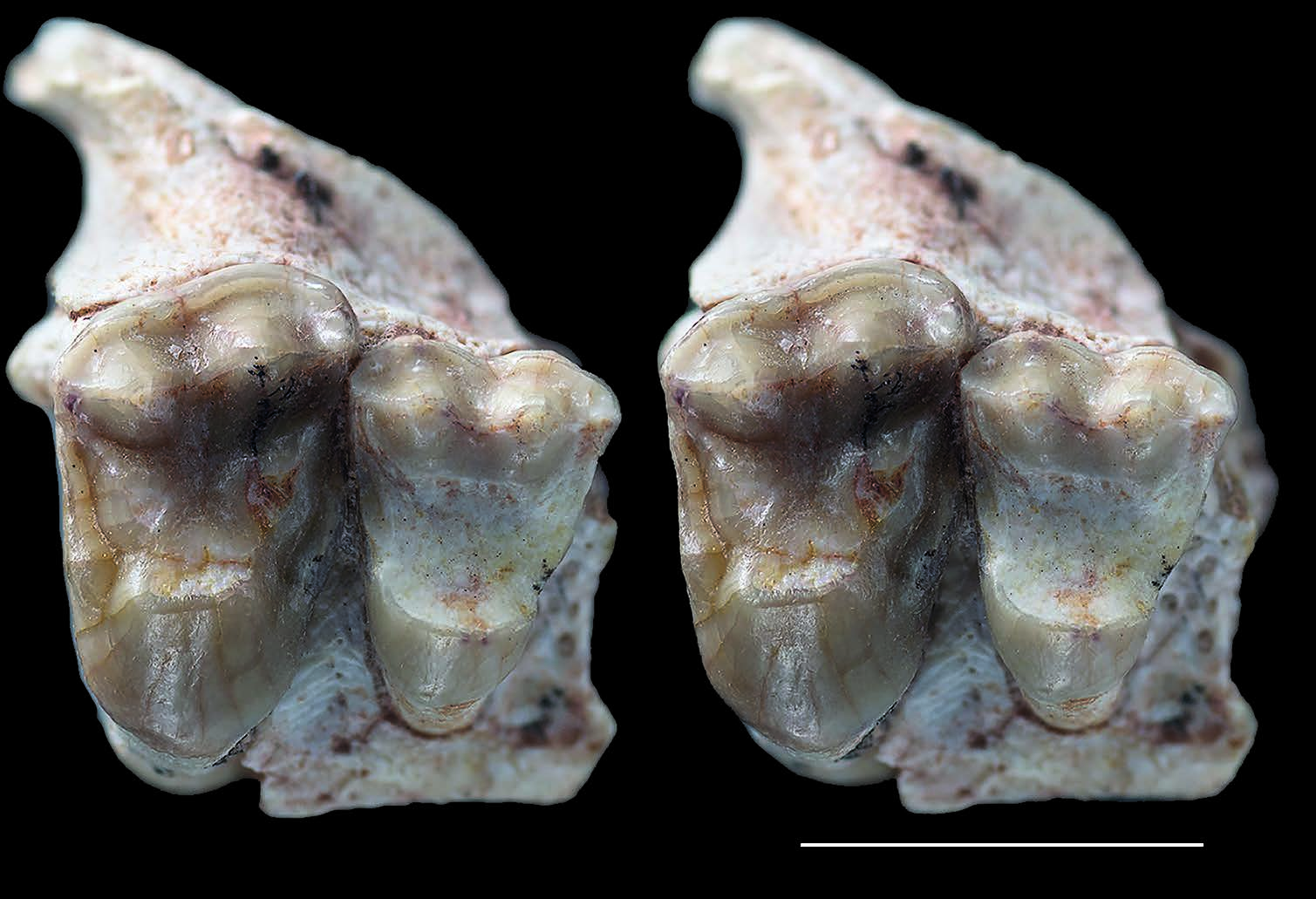
A well-preserved lateral ascending process of a right maxilla (MHNC 13868) is referred to Simoclaenus sylvaticus . This specimen bears the root of the canine and the four premolars, which were unknown for this taxon ( Fig. 8 View FIG ). The upper premolars of MHNC 13868 are only slightly larger than those of Molinodus suarezi described by Muizon & Cifelli (2000: 59), but this difference could be interpreted as related to individual variation in size. The major criterion that convinced us to relate this specimen to S. sylvaticus is the size of the canine. Although the crown of that tooth is not preserved the section of the root at the level of the alveolar border is relevant to its size. It is noteworthy that no upper canine is known for M. suarezi . We therefore compared the section of the upper canine of MHNC 13868 to that of the lower canine of MHNC 13883. Because the Tiupampa kollpaniines are relatively unspecialized, it is hypothesized that the width and length (at the level of the alveolar plane) of the upper and lower canines did not differ significantly even if generally the upper canines are slightly larger than the lowers. This remains an assumption, as no kollpaniine has ever been documented by both upper and lower canines.
Here, the upper canine of MHNC 13868 ( Simoclaenus ) is 87% longer and 55% wider than the lower canine of MHNC 13883 ( Molinodus ) ( Table 3). Even taking into account the approximation in comparing upper and lower canines, such a size difference is highly significant and we regard as unlikely that the two canines could belong to the same taxon of kollpaniine. We therefore, tentatively refer MHNC 13868 to S. sylvaticus rather than to M. suarezi . Furthermore, the size difference between the P4 and M1 of the holotype of S. sylvaticus and referred specimen described here could be related to sexual dimorphism in this species as evoked above.
The premolars are well preserved and bear very reduced dental wear. The size of P3 and P4 is slightly larger than the equivalent teeth in Molinodus suarezi , which are known on one specimen only (MHNC 1247) (see Muizon & Cifelli 2000: 58) ( Table 4). P1 and P2 are unknown for M. suarezi . Although this size difference is moderate (c. 10%), because of the large size of the canine, we rather refer this specimen to Simoclaenus sylvaticus . Furthermore, the anteroposterior length of the upper premolar row of MHNC 13868 (11.31 mm) is only slightly smaller than the lower premolar row of the holotype of S. sylvaticus (11.97 mm), on which the alveoli of p1-3 and the p4 are preserved. Therefore, the mesiodistal length of the upper premolar row of the new specimen (MHNC 13868) correctly matches the length of the lower premolars row of the holotype (MHNC 8348).
P1 is peg-like, single-rooted, and compressed transversely. It is implanted vertically in the maxilla as is observed on the alveolus of the p1 of the holotype (MHNC 8348). This condition suggests a relative shortness of the rostrum and is congruent with the anteroposterior compression of the lower cheek teeth observed on the mandible. It is noteworthy that the p1 of Molinodus described above (MHNC 13883) differs from the condition observed in Simoclaenus in being slightly procumbent, thus suggesting a more elongated rostrum. As preserved, the P1 of Simoclaenus on MHNC 13868 is longer than high and no significant wear facet can be observed at the apex of its crown. In lateral view the tooth is roughly symmetrical anteroposteriorly. A small diastema separates P1 from the canine anteriorly and from P2 posteriorly. P2 is triangular in occlusal view and bears three roots. It is longer than wide. It presents a conspicuous inflation on its distolingual edge, which can be regarded as an incipiently developed protocone (protoconal bulge). The paracone forms most of the tooth and is as high as long. Its mesial edge is wide and blunt whereas its distal edge is thin and forms a sharp crest. The tooth bears no cingulum but a small cusp at the distolabial edge of the paracone could be regarded as a metastyle. This style contacts the anterior edge of P3 and no diastema separates the two teeth. P3 is slightly wider than long, triangular in occlusal view, and bears three roots. The mesial and distal edges are markedly concave, a condition which individualizes a well-developed protocone lingually. This cusp is approximately two thirds the height of the paracone. It is as long as wide. It has a very convex lingual edge and a flat labial aspect. These two edges of the tooth are separated by sharp pre- and postprotocristae, which join the base of the crown at the level of the greatest concavity of the mesial and distal edges of the tooth. Labially the paracone is approximately twice as long as wide. Its mesial edge bears a smooth crest and its distal crest is slightly obliterated by an elongated wear facet, which extends from the apex of the tooth to the posterior base of the paracone. At the mesial angle of the paracone is a marked parastyle. From this cusp, a conspicuous cingulum extends on labial edge of the paracone. At the distal end of the cingulum is a small metastyle. P4 has a pattern similar to that of P3 but it is mesiodistally shorter and transversely wider. Its mesial and distal edges are slightly concave but to a much lesser extent than the condition observed on P3. The paracone is smaller (in height and volume) and the protocone is more voluminous than on the preceding tooth. The pre- and postprotocristae are more developed than on P3 and extend on the anterior and posterior edges of the paracone. The postprotocrista even joins the distolabial angle of the tooth and contacts the postparacrista. The latter bears a narrow wear facet on its mesial two thirds only. The distal end of the postparacrista bears, on its lingual aspect, a hint of inflation, which can be interpreted as an incipient metacone. A tiny inflation on the postprotocrista may also be regarded as an incipient metaconule. On the distal edge of the protocone, is a distinct postcingulum. On its mesial edge the precingulum is weakly developed. On the mesiolabial angle of the tooth the parastyle is more developed than on P3. From this style, a well-developed labial cingulum extends distally until the metastylar angle of P4.
Little is preserved of the palatal process of the maxilla, but the lateral wall of the rostrum from the anterior edge of the canine to the anterior root of the zygomatic arch is relatively complete and all its edges likely correspond to sutures with adjacent bones except for a small U-shaped-break in the posterodorsal angle ( Fig. 8C View FIG ). The anterodorsal edge of the specimen probably corresponds to the nasal-maxilla suture. Approximately above the P1-P2 embrasure, the edge of the maxilla distinctly protrudes medially. Anterior to this indentation, the suture is apparently anteroposteriorly oriented or slightly oriented anterolaterally. Posterior to it, the suture diverges posterolaterally, thus indicating a widening of the nasals posteriorly, as generally observed in early diverging metatherians and eutherians (e.g., deltatheroidans, pucadelphydans, Zalambdalestes, Kulbeckia, Alcidedorbignya). On the posterior limit of the maxilla, as preserved, is a deep groove probably for the articulation of the anterior process of the jugal. This groove (maxillajugal suture) approximately corresponds to the level of the anterior edge of the orbit and the posterior limit of the rostrum, which corresponds to the posterior edge of P4. On the posterodorsal angle of the specimen, in lateral view, is a distinct small notch (dorsal to the breakage notch mentioned above), which was receiving the anterolateral angle of the frontal. If this interpretation is correct, the nasal and lacrimal of Simoclaenus were distinctly separated, which likely represents a crown Placentalia condition (frontal-maxilla suture present) ( Muizon et al. 2015). The posterior edge of the maxilla, between this notch and the dorsal end of the jugal groove, likely corresponds to the lacrimal-maxilla suture. Anteriorly, several grooves mark the edge of the maxilla immediately anterior to the canine, which we interpret as the premaxilla-maxilla suture. This suture is almost straight, slightly concave anteriorly, and sub-vertical, forming an angle of approximately 95° with the alveolar plane ( Fig. 8C View FIG ).
The lateral aspect of the maxilla is markedly elevated and erected, to a greater extent than the condition observed in the pantodont Alcidedorbignya inopinata from the same locality ( Muizon et al. 2015). This condition apparently resembles that of Baioconodon nordicum (YPM-PU 14234) from the earliest Palaeocene (Puercan) of Mantua lentil of Wyoming.
The anterior opening of the infraorbital canal is 2.08 mm high and located above the mesiolabial root of P3. In this respect Simoclaenus differs from the condition in Maiorana and Baioconodon, in which the infraorbital foramen is located more posteriorly, above the mesiolabial root of P4. A condition similar to that of Maiorana and Baioconodon is observed in Didolodus multicuspis (MACN 10690), in which the infraorbital foramen is located above the distal edge of P3 and most of the P4 (Gelfo, personal communication). Anterior to this foramen is a tiny opening located above the anterior root of P2, and which probably represents a nutrient foramen. In its anterior region, the maxilla is not inflated laterally by the canine as is observed in Alcidedorbignya, due to the fact that the canine was compressed transversely. This condition particularly resembles the “condylarths” of Mantua lentil of Wyoming, Baioconodon and Maiorana.
The third specimen referred to Simoclaenus sylvaticus is a partial maxilla with M1-2 (MHNC 13876), ( Fig. 9 View FIG ). The M2 perfectly matches the size and morphology of that of MHNC 8348 described by Muizon & Cifelli (2000). The small differences between the two teeth are the presence, in MHNC 13876, of a slightly larger parastyle, a cuspule on the preparaconular crista located between the paraconule and the parastyle and a slight inflation of the lingual end of the precingulum (cuspule?). The M2 of MHNC 13876 has the characteristic asymmetrical morphology observed in Molinodus and Simoclaenus , in which the paracone is distinctly more labial than the metacone ( Muizon & Cifelli 2000: fig. 2A-C, fig. 14C). This condition is not observed on the M1 of Molinodus , the paracone of which is not shifted labially. The most interesting characteristic of this specimen is in the relative wear stage of the M2 and M1. The M2 has a wear stage approximately similar to that of the M2 of MHNC 8348. In contrast the M1 is totally excavated in its lingual two thirds: the protocone, the protocristae and the conules, the pre-and postcingula, and the trigon basin have disappeared; the only preserved elements of the tooth are the para- and metacones, the styles and the labial cingulum. This wear stage is clearly more advanced than that observed on the M1 of MHNC 8348, in which the protocone and conules are coalescent but still identifiable, the protocristae and the pre-and postcingula are distinctly observable. This stage of wear of M2-1 of MHNC 8348 is comparable to that observed on the maxilla of Molinodus suarezi (MHNC 1247) described by Muizon & Cifelli (2000). In fact, the extensive wear of the M1 of MHNC 13876 is comparable to the condition that could be observed on a DP4, an interpretation impossible given the fact that the posterior tooth is undoubtedly an M2, which presents the characteristic asymmetrical morphology of that tooth ( Muizon & Cifelli 2000). Furthermore, the roots of the M1 are long and the lingual root perforates the floor of the orbit, which suggests that it is not a deciduous molar.
In ventral view, laterally to M2, the maxillar process of the zygoma and the anterior border of the orbitotemporal fossa are preserved. The latter is at the level of the apex of the metacone of M2, while it is at the level of the paracone of M3 on the maxilla (MHNC 8348) referred to Simoclaenus by Muizon & Cifelli (2000). In Molinodus the anterior edge of the orbitotemporal fossa is at the level of the paracone of M3 and in Baioconodon (YPM-PU 14234) and Maiorana (YPM-PU 16667 and 14171) it is at the level of the metacone of M3. However, this condition is likely to be individually variable since in Alcidedorbignya inopinata out of 13 specimens, in which it can be observed, three of them (see Muizon et al. 2015: fig. 10B) have an anterior edge of the orbitotemporal fossa at the level of the metacone of M2, as observed in the Simoclaenus specimen described here. In the other ten specimens, the anterior edge of the fossa is at the level of the paracone of M3. In the Alcidedorbignya sample the anterior position of M2 is observed in young adults and may be related to the ontogenetic increase of the length of the rostrum.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.