
Uroplatus finiavana, Ratsoavina, Fanomezana Mihaja, Louis, Edward E., Crottini, Angelica, Randrianiaina, Roger-Daniel, Glaw, Frank & Vences, Miguel, 2011
|
publication ID |
https://doi.org/ 10.5281/zenodo.206497 |
|
DOI |
https://doi.org/10.5281/zenodo.6183301 |
|
persistent identifier |
https://treatment.plazi.org/id/039EBC42-FF80-FFE8-FF26-3F9B0536FB72 |
|
treatment provided by |
Plazi |
|
scientific name |
Uroplatus finiavana |
| status |
sp. nov. |
Uroplatus finiavana sp. nov.
Holotype. ZSM 328/2004 ( FGZC 625, adult male, hemipenes partly everted) collected at Montagne d'Ambre, 700–1000 m above sea level (a. s. l.) on 19–23 February 2004 by F. Glaw, M. Puente, R.D. Randrianiaina & A. Razafimanantsoa.
Paratypes. ZSM 1132/2003 (FG/MV 2002-2387, adult male); ZSM 1133/2003 (FG/MV 2002-2388, adult male); ZSM 1134/2003 (FG/MV 2002-2390, adult female); and ZSM 1135/2003 (FG/MV 2002-3083, adult male), all collected at Montagne d'Ambre, on 17–20 February 2003 by F. Glaw, R. D. Randrianiaina & A. Razafimanantsoa.
ZSM 322/2004 ( FGZC 619, adult female), ZSM 323/2004 ( FGZC 620, adult female), ZSM 324/2004 ( FGZC 621, adult female), ZSM 325/2004 ( FGZC 622, adult female), ZSM 326/2004 ( FGZC 623, adult male), ZSM 327/ 2004 ( FGZC 624, adult male), ZSM 329/2004 ( FGZC 626, adult male), all collected at Montagne d'Ambre, 700– 1000 m a. s. l. on 19–23 February 2004 by F. Glaw, M. Puente, R. D. Randrianiaina & A. Razafimanantsoa.
Diagnosis. Uroplatus finiavana sp. nov. differs from all other taxa of the U. fimbriatus species group ( U. fimbriatus , U. giganteus , U. henkeli , U. sikorae and U. sameiti ) and U. lineatus by its smaller size (adult SVL 52–65 mm versus at least 85 mm) and lack of lateral membranous fringes on any part of the body and limbs; and from Uroplatus alluaudi , U. guentheri , U. pietschmanni , and U. malahelo by its smaller size (adult SVL 52–65 mm versus 69–81 mm), laterally compressed body with a vertebral keel, and more triangular head.
The new species is most similar to the other species of the Uroplatus ebenaui group ( U. ebenaui , U. phantasticus and U. malama ).
Uroplatus finiavana differs from U. malama by its smaller body size (SVL 52–65 mm versus 71–77 mm); shorter (TaL/SVL 0.42–0.65 versus 0.61–0.72) and narrower tail (TaW/SVL 0.05–0.14 versus 0.18–0.24), an unpigmented oral mucosa (versus blackish oral mucosa), and the presence of spines on the body, head, limbs and tail base (versus their absence).
Uroplatus finiavana differs from U. ebenaui by its slightly larger body size (SVL 52–65 mm, mean 58.2 mm versus 45–63 mm, mean 54.3 mm), longer tail (TaL/SVL 0.42–0.65 versus 0.22–0.40), longer forelimb (ForL/SVL mean 0.42 versus 0.38), posteriorly curved interorbital ridge (versus almost straight), the median posterior extension of the ridge (1.6–3.0 mm versus 0.0– 1.1 mm), more extended neck triangle (neck-triangle length 7.3–14.2 mm versus 1.7–3.4 mm), and by its non-pigmented oral mucosa (versus blackish oral mucosa).
Uroplatus finiavana differs from U. phantasticus in its slightly smaller body size (SVL 52–65 mm, mean 58.2 mm versus 52–76 mm, mean 60.3 mm), shorter (TaL/SVL 0.42–0.65 versus 0.62–0.76) and narrower tail (TaW/ SVL 0.05–0.14 versus 0.16–0.20), and its unpigmented oral mucosa (versus blackish oral mucosa).
In addition, U. finiavana differs from all other species of the Uroplatus ebenaui group (see Figures 6 View FIGURE 6 and 7 View FIGURE 7 ) and from all other species of Uroplatus ( Greenbaum et al. 2007; Raxworthy et al. 2008) by its substantial degree of genetic differentiation.
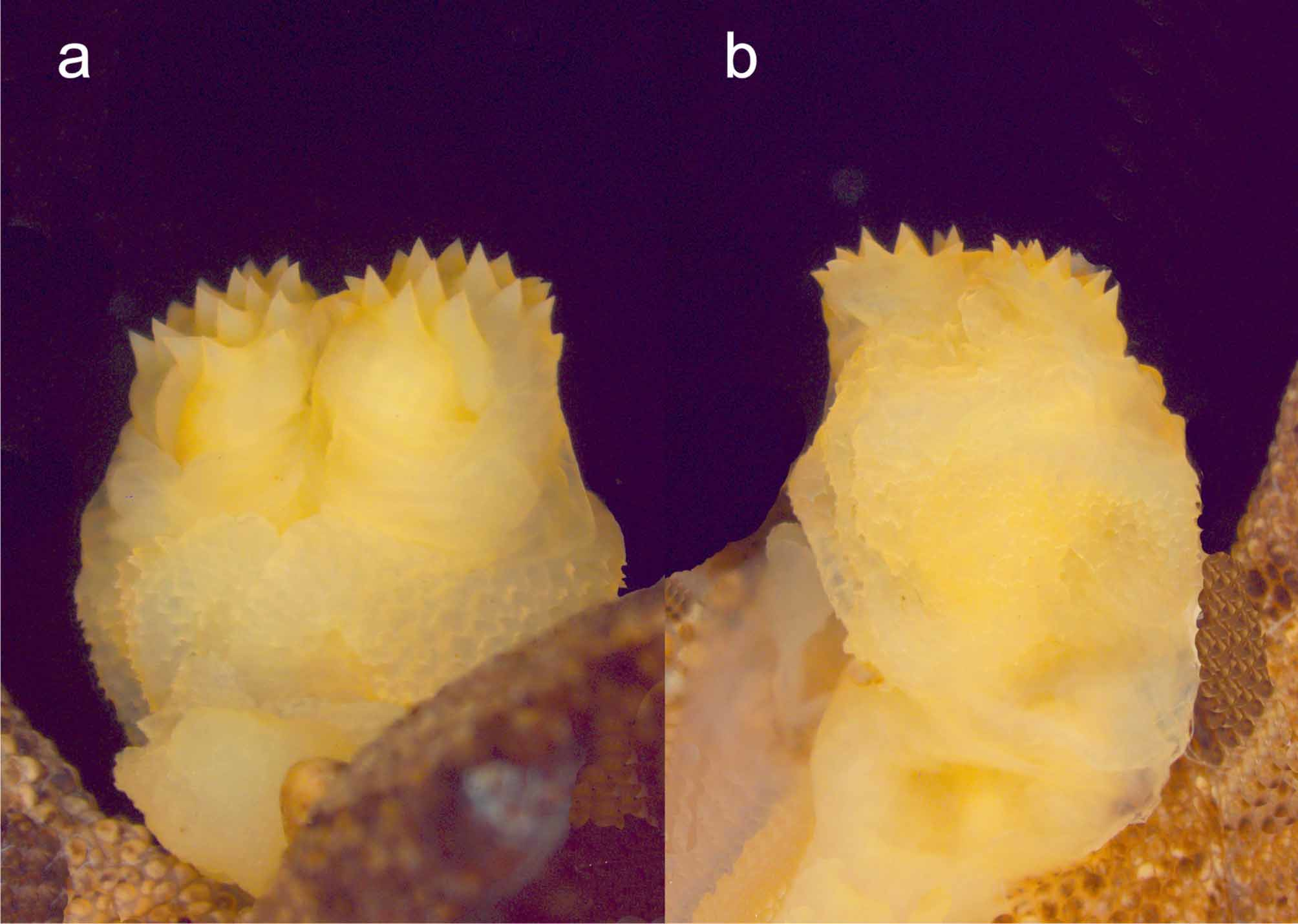
Description of the holotype. Male specimen in good condition, with original tail attached to the body and partially everted hemipenes. Measurements and counts of the holotype: snout-vent length 59.0 mm, tail length 35.3 mm, maximum tail width 5.7 mm, and right forelimb length 25.5 mm. Head triangular in dorsal view, postorbital region (measured from posterior border of eye to anterior border of ear opening) of similar length to that of the snout (from anterior eye border to snout tip); snout sloping strongly and continuously downward anteriorly; snout depressed, short (1.3 times longer than eye diameter); canthus rostralis indistinct; eyes large, bulging slightly above dorsal surface of cranium, directed laterally, pupil vertical with crenate borders; ear opening very small (horizontal diameter 0.6–0.8 mm), its opening facing posterolaterally, but also posteroventrally (ear opening clearly visible in ventral view); nares laterally oriented; body somewhat laterally compressed, without lateral fringes; limbs well developed, without fringes, forelimb reaches beyond tip of snout and almost to the groin (forelimb length/axillagroin distance 98%), hind limb reaches beyond axilla (hind limb length/axilla-groin distance 132%); tail 60% of snout-vent length, membranous borders of the tail narrow (maximum width on each side 1.6 mm) and completely absent from the distal tip of the tail. Nares separated from each other by eight small granular scales, from the first supralabial by one scale, and from the rostral scale by two scales; first supralabial taller than others; rostral entire, much wider than tall; mental scale very small, not differentiated from infralabial scales (total series of infralabials, left and right, plus the intervening mental scale yields a count of 35); no enlarged postmental scales or chin shields; dorsal and ventral scales of head, neck, body, limbs, and tail small, granular, juxtaposed and largely uniform in size, except for the irregular lines on the head and body which consists of series of slightly enlarged scales. Two curved lines starting at the posterolateral parts of the head converge in the neck forming a V-shaped pattern (neck triangular line). A curved, moderately distinct and posteriorly directed line is present between the eyes and connects the supraciliary spines. Several spines on the posterior parts of the head, on hind limbs and a single pointed flap on the posterior portion of each upper eyelid; upper eyelid becomes broader as it approaches the parietal region of the head.
Coloration: All dorsal surfaces are brown in color except the tail which is light brown mottled with greybrown and with few dark spots on the neck and one larger dark spot on the middle of the back after more than 6 years in alcohol. The mottling of the body is configured as a series of fine posteriorly directed markings along the dorsum. Tail is uniformly light brown. Two whitish spots present below each eye. Chin and throat beige with a distinct blackish wide V-marking that merges together to form an indistinct dark longitudinal line that fades on the throat. Venter is light grey with few scattered small darker spots. Lower hind limbs and feet are slightly darker than other ventral surfaces. Postpygal portion of tail is marked with a whitish spot. Oral mucosa unpigmented.
Variation. The dorsal ground color varies from light beige (ZSM 323/2004, 325/2004, 326/2004, 327/2004) to reddish-brown (ZSM 324/2004), to dark-brown (ZSM 322/2004) and grey (ZSM 329/2004). Dark spots are most evident in those paratypes with light beige ground color, whereas the other specimens are largely uniform in coloration and lack prominent dark spots.
The ventral side of all paratypes is similar to that of the holotype except for two characters. First, the V-marking on the chin of the holotype is absent from two of the paratypes (ZSM 322/2004 and ZSM 329/2004) or is indistinct (ZSM 1135/2003 and ZSM 1132/2003) and also, the scattered dark spots are absent from three paratypes (ZSM 324/2004, ZSM 322/2004 and ZSM 329/2004). One of the paratypes (ZSM 327/2004) shows distinct lateral dark line starting slightly under the axilla and continues to the groin. The presence of a white spot in the postpygal area is not constant and is only found in three of the eleven paratypes.
Color photographs most likely showing Uroplatus finiavana have appeared in several publications, including those of Andreone (1991), Glaw & Vences (1994: color photo 252), Böhme & Henkel (1995: Figures 11, 12, 13, 14), Svatek & van Duin (2002), Glaw & Vences (2007: 378) and Schönecker (2008: 125–126). The same pattern as those encountered in the preserved specimens is observed in living animals. In some specimens, the markings on the head and the dorsum are underlined with darker coloration. Whitish spots below each eye are sometimes absent or are reduced to a single one.
Hemipenis structure. Because the holotype does not have a fully extruded hemipenis, we used one of the paratypes ZSM 1133/2003 with fully everted hemipenis for the description ( Figure 4 View FIGURE 4 ). In common with the other species of the ebenaui group, the hemipenis bears two lobes that become more obvious towards the apical region. The calyx displays a protuberance with a honeycomb appearance within which the sulcus spermaticus is concealed. The latter has a smooth surface. One to four folds separate the protuberance from the apex, which exhibits several papillae that wrap a single structured pedunculus in each side. A more in-depth description and comparisons with other taxa will be presented elsewhere.
Etymology. The specific name is derived from the Malagasy word " finiavana " meaning initiative. We refer to the fact that we took the initiative to name the species following years of this taxon having been recognized as likely being distinct. The name is used as a noun in apposition.
Distribution. In several herpetological surveys carried out in 1994, 2000, 2003, 2004, 2006 and 2009, the new species was encountered in the rainforest of Montagne d’Ambre National Park between 700–1350 m a.s.l. and in secondary forest fragments of lower elevation close to the town of Joffreville, where both U. finiavana and U. ebenaui occur sympatrically. This protected area is completely isolated from the major rainforest blocks of northern and eastern Madagascar and covers a surface area of 182 km 2 ( Nicoll & Langrand 1989; Raxworthy & Nussbaum 1994). D’Cruze et al. (2008) reported encountering this new species in the adjacent forest (Réserve Spéciale Forêt d’Ambre) that has a lower elevation range of 400–850 m a.s.l.; but we cannot exclude the possibility that these records may partly, or entirely, refer to U. ebenaui . Records of U. ebenaui from the neighboring rainforest blocks (Anjanaharibe-Sud, Manongarivo, Marojejy, Tsaratanana and Makira) might be referable, by DNA sequences and morphology, to other taxa.
Habitat and habits. The holotype was collected in primary forest in Montagne d’Ambre National Park during a night survey. The species is nocturnal, arboreal and is remarkably common within the national park, especially around 800–900 m a. s. l., where it is possible to encounter 10 individuals during a single night’s walk (for instance on the path “Voie des mille arbres”). Remarkably, most of the encountered specimens have lost their original tail. One individual was observed feeding on a cockroach.
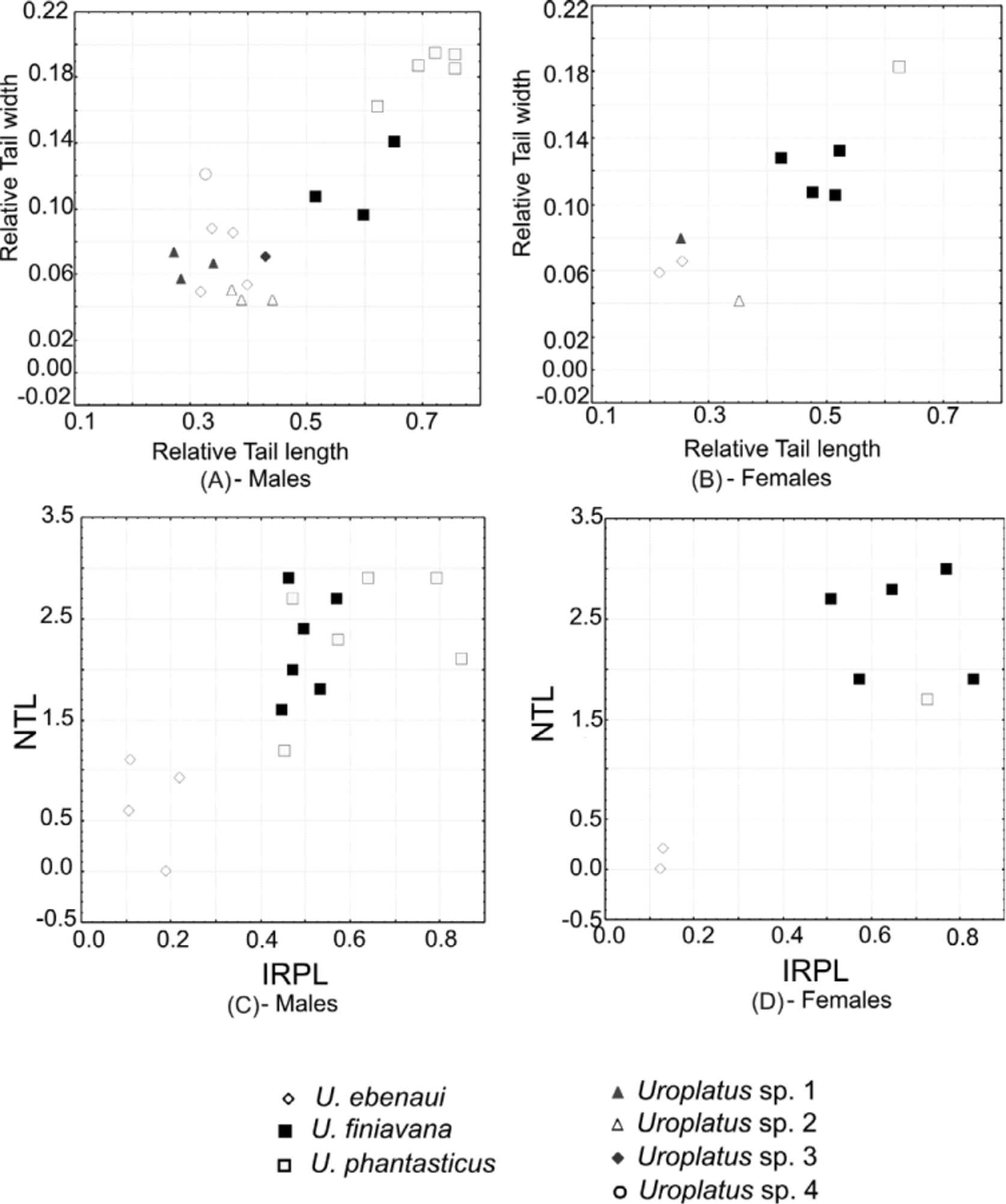
Morphometric differentiation. Relative tail length and relative tail width were recorded as ratios relative to SVL; these values were calculated separately for males and females. Figures 5 View FIGURE 5 a and 5b show that the same pattern is evident in both sexes. The new species, U. finiavana , shows intermediate values of tail length and tail width relative to two other described species of its clade ( U. ebenaui and U. phantasticus ), and the differences are statistically supported (Mann-Whitney U test, pooling males and females; P <0.01). Figures 5 View FIGURE 5 c and 5d compare neck triangle (NTL) and medial posterior projection of the interorbital ridge length (IRPL), revealing only two clusters; U. ebenaui differs from U. finiavana and U. phantasticus in its smaller values for these two parameters. Significant differences only exist between U. ebenaui and U. finiavana for NTL, IRPL and relative ForL (P <0.01). There is no significant differentiation among the three species in relative hindlimb length.
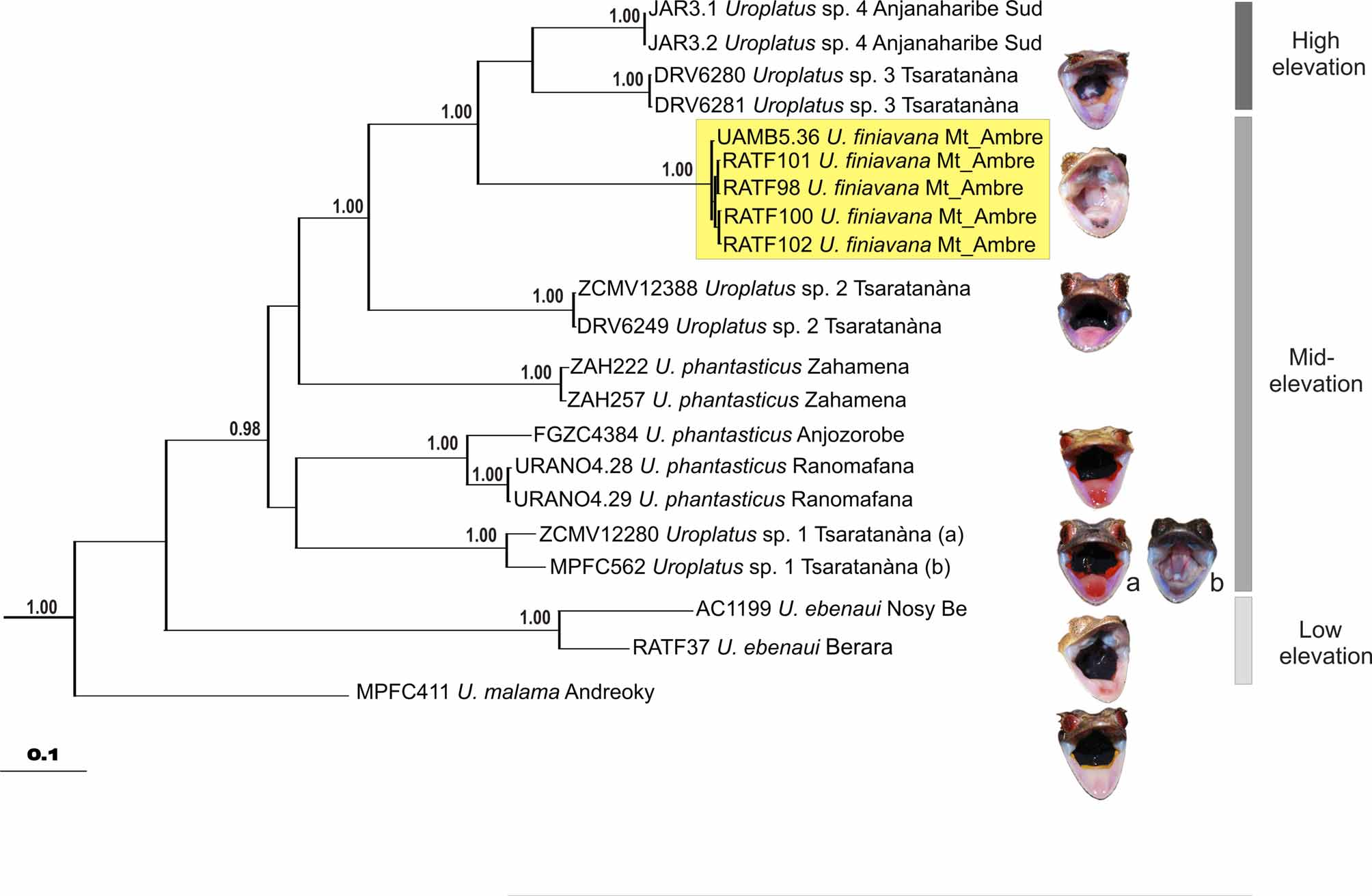
Molecular differentiation and phylogenetic relationships. The phylogenetic tree inferred from the Bayesian analysis of the mitochondrial ND4 gene is shown in Figure 6 View FIGURE 6 . We emphasize that this tree should be considered as preliminary as it is based on comparatively short sequences of a single gene. Rather than a reliable phylogeny, this tree wants to depict genetic similarities and differences among the specimens studied. On this basis, the morphological differences observed between the new species and U. ebenaui from the type locality ( Nosy Be) and U. phantasticus are supported by molecular data. The tree displays a subdivision of the U. ebenaui group into seven deeply divergent clades, with U. malama occupying a basal position. Besides the described species, the tree contains additional deep genealogical lineages, which we here classify as candidate species according to the scheme of Vieites et al. (2009). First, based on our initial assessment of identity of the nominal species, we assume that four of these are included in the phylogram: U. ebenaui , U. phantasticus , and U. malama , plus U. finiavana sp. nov. as described herein. Secondly, Uroplatus sp. 4 from Anjanaharibe-Sud is an Unconfirmed Candidate Species (UCS) due to its high genetic divergence but insufficient morphological data to assess its status. Uroplatus sp. 1, 2 and 3 can be considered as Confirmed Candidate Species (CCS) because of their concordant divergence in both molecular and morphological data (e.g.: tail shape) (see Figs. 5 View FIGURE 5 and 8 View FIGURE 8 ).
Uroplatus finiavana is the sister species of the clade consisting of high elevation CCS Uroplatus sp. 3–4, and this sister relationship is strongly supported [Bayesian Posterior Probability (BPP)= 1.00]. The cluster of U. finiavana and Uroplatus sp. 3 and 4 is the sister clade of Uroplatus sp. 2 (strongly supported relationship: BPP= 1.00).
The analyzed samples of U. phantasticus do not cluster as a monophyletic group. Uroplatus phantasticus from Anjozorobe and Ranomafana constitutes the sister species of U. sp. 1, although this sister-relationship is not strongly supported, and U. phantasticus from Zahamena is basal to the clade composed of U. finiavana and U. sp. 2–4, but also in this case the phylogenetic relationships are not reliably resolved. The monophyly of the clade consisting of U. finiavana , U. sp. 1–4 and U. phantasticus from Zahamena, Anjozorobe and Ranomafana is strongly supported (BPP= 0.98). The phylogenetic tree is somewhat concordant with altitudinal range data: U. ebenaui from the lowland occupies a basal position, followed by the CCS of U. ebenaui , both U. phantasticus lineages and U. finiavana from mid-elevations, and by the UCS U. sp. 4 and the CCS U. sp. 3 from the high mountains in a deeply nested position.
Among the analyzed taxa, exceptionally large genetic distances (uncorrected pairwise distances shown in table 2) in the ND4 gene are evident. The highest observed value is 37.5% and is observed between U. ebenaui from Nosy Be and U. phantasticus from Ranomafana (individual URANO4.29), whereas the smallest value among major clades is found between the candidate species U. sp. 3 and U. sp. 4.
Uroplatus finiavana is highly divergent from all other lineages of the Uroplatus ebenaui species complex. The uncorrected pairwise genetic distances of U. finiavana from U. sp. 1–4, U. phantasticus from Zahamena, U. phantasticus from Ranomafana and Anjozorobe, U. ebenaui and U. malama ranges from 20.8 to 34.5%. These large genetic distances corroborate the separation of the newly described species from all other lineages and known species included in this study. Moreover, relatively large genetic divergences are found between individuals assigned to the same species. For instance, the uncorrected genetic distance between U. phantasticus from Ranomafana and Anjozorobe is 6.6% and the uncorrected genetic distance between U. ebenaui from Nosy Be and Berara is 18.2%.
As our goal was not a detailed phylogenetic reconstruction but a test for concordant differentiation in two independent genetic markers for integrative taxonomic purposes (see Padial et al. 2010), we analyzed the nuclear Cmos sequences separately from the mitochondrial ND4 sequences, and used a network analysis to better visualize the low degree of haplotype differentiation among the focal taxa. The haplotype network reconstruction based on the C-mos data revealed no evidence of haplotype sharing between U. finiavana and other species and candidate species of the U. ebenaui group ( Figure 7 View FIGURE 7 ). This congruence between mitochondrial and nuclear data supports the status of the new species as a separate evolutionary lineage and indicates the likely absence of gene flow among different lineages of the U. ebenaui group. Individuals from Tsaratanana assignable to Uroplatus sp. 1–3 in the haplotype network ( Figure 7 View FIGURE 7 ) are in grey and haplotype sharing among some of these candidate species occurs. For instance, Uroplatus sp. 2 from Tsaratanana (ca 2000 m a. s. l.), Marojejy and Makira’s western slope have the same haplotype as Uroplatus sp. 3 from the Andrevorevo part of the Tsaratanana massif (ca 1700 m a. s. l.).


| ZSM |
Bavarian State Collection of Zoology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.