
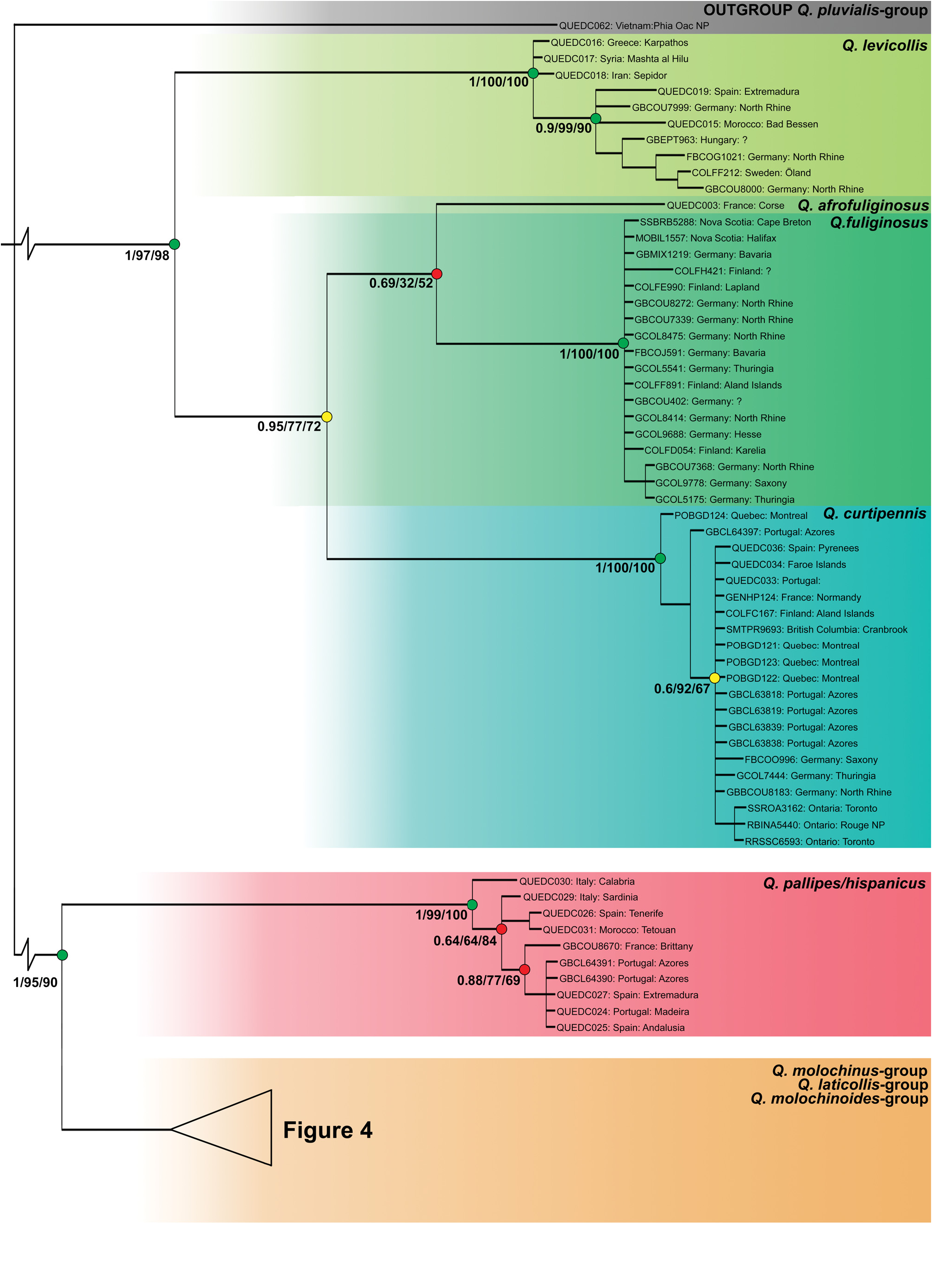
Quedius hispanicus Bernhauer, 1898
|
publication ID |
https://doi.org/ 10.37520/aemnp.2022.017 |
|
publication LSID |
lsid:zoobank.org:pub:28D55112-98B1-49A5-B382-58B1B068570B |
|
DOI |
https://doi.org/10.5281/zenodo.7503703 |
|
persistent identifier |
https://treatment.plazi.org/id/038987A0-FFD0-4B35-B388-A4FE1015F9EA |
|
treatment provided by |
Felipe |
|
scientific name |
Quedius hispanicus Bernhauer, 1898 |
| status |
|
Quedius hispanicus Bernhauer, 1898 View in CoL
( Figs 1 View Fig , 2D View Fig , 3 View Fig , 9A,B View Fig , 14C View Fig , 19 View Fig )
Quedius hispanicus Bernhauer, 1898: 341 View in CoL [Type locality: Cartagena und Cadix in Spanien] stat. rev.
Quedius cobosi Coiffait, 1964: 293 View in CoL [Type locality: Espagne: Province de Almeria, Seron], syn. nov.
Quedius hammianus Sharp, 1911:57 View in CoL [Type locality:Anglia:Deal, Strood, Lymington, and Hayling Island], syn. rev.
Quedius rufulus Blümml, 1898: 216 View in CoL [Type locality: Spanien, Granada], syn. rev.
Quedius secundus Last, 1952 View in CoL [Type locality:Studland, Dorset; Mudeford, Hampshire; Chichester], syn. rev.
References. Fൺඎඏൾඅ (1865): 16 (synonym of Q. molochinus View in CoL ); Kඋൺൺඍඓ (1867): 414 (synonym of Q. molochinus View in CoL ); GൺඇGඅൻൺඎൾඋ (1895): 403 (synonym of Q. molochinus View in CoL ); BൾඋඇHൺඎൾඋ (1900): 46 (distribution); Pඈඋඍൺ (1907): 129 (synonym of Q. molochinus View in CoL ); Gඋංൽൾඅඅං (1924): 81 (subspecies of Q. molochinus View in CoL ; characters); SർHൾൾඋඉൾඅඍඓ (1933): 1457 (synonym of Q. picipennis View in CoL ); MඣඊඎංGඇඈඇ (1937): 3 (as Q. molochinus pallipes View in CoL ); KඈඋGൾ (1960b):69 (synonym of Q. pallipes View in CoL ); Cඈංൿൿൺංඍ (1961): 56 (characters and distribution); (1978):196 (characters); Hൺඇඌൾඇ (1962): 319 (characters and distribution); Sආൾඍൺඇൺ (1962a): 134 (characters); (1963): 39; (1970): 61 (distribution); Lൺඌඍ (1963): 45 (synonym of Q. simplicifrons View in CoL ); LඈHඌൾ (1964): 211 (characters and distribution); Hඈඋං- ඈඇ (1965): 276 (distribution); Bඈඋൽඈඇං (1976a): 100 (synonym of Q. pallipes View in CoL ); Pඈඉൾ (1977): 31 (distribution); LඎർHඍ (1987): 109 (distribution); LඈHඌൾ & LඎർHඍ (1989): 172 (distribution); Iඌඋൺൾඅඌඈඇ (1990): 3 (distribution); BඈඋGൾඌ (1990): Table II (distribution); WൾඅർH (1993): 229 (ovariole number and ovary structure); Oඎඍൾඋൾඅඈ et al. (1995): 80 (distribution; as Q. pallipes View in CoL ).
Type material examined. Quedius cobosi : Pൺඋൺඍඒඉൾ: J ( MNHN), labelled: “ PARATYPE [red label] / Quedius cobosi Coiff. H. COIFFAIT det. 1964 / Laujar Almeria - España A. Cobos coll.”
Additional material examined. DENMARK: Lakolk, [55.14, 8.49], debris on beach, 1.XI.2013, leg. J. Pedersen & A. Brunke (1 J NHMD); Skallingen, [55.50, 8.25], 17.X.1959, leg. F. Bangsholt (2 JJ NHMD). GERMANY: Archsum, [Sylt], [54.87, 8.39], IX.1901 (1 SDEI); Insel Borkum, [53.58, 6.66], 12.VIII.1937, leg. Struse (2 ♀♀ 4 JJ NMW); N Dagebüll, Nordfriesland, 54.7361, 8.7094, salt pasture among stones and clay on dam, 25.VIII.2017, leg. Wrase & Laser (1 J cSch); Memmert, Juist, [53.64, 6.88], 13.VIII.1917, leg. Alfrum (1 SDEI). FRANCE: Bordeuax, St. Louis, [44.94, -0.52], 22.II.1964 leg. C. Jeanne (1 JcBor); Camargue, Etang du Fangassier, [43.42, 4.63], 20.V.1994 salt marsh, leg. Schülke & Grünberg (1 J cSch); Sainte Marie la Mer, [42.72, 3.03], 28.IV.1955, leg. J.T. Skovgaard (1 ♀ NHMD). PORTUGAL: Lissabon, [38.76, -8.87], leg. Von Heyden (1 J 2 ♀♀ SDEI); Lusitania, Lissabon, [38.76, -8.87], IV.-V.1910, leg.A. Schartzmayr (1 J 1♀ SDEI). Aඓඈඋൾඌ: Faial.Baia da Areia das Fontes 1km N of Praia do Norte, [38.61, -28.74], under stones, grassy ground, near rocky shore, 2.IV.1957, leg.Brinck and Dahl (5 ♀♀ MZLU); Faial, Caldeira, [38.58, -28.71], 10.IV.1957. leg. Brinck and Dahl (3♀♀ MZLU); Faial, Horta, Porto Pim., [38.52, -28.62], sandy ground, 7.IV.1957 leg.Brinck and Dahl (1 ♀ MZLU); Faial. Nasce Agua 3 km S of Cedros, [38.60, -28.68], ravine, under stones, 5.IV.1957, leg. Brinck and Dahl (1 J MZLU); Faial, Praia do Almoxarife, [38.55, -28.61], sandy grassy ground, under stones, 31.III.1957. leg. Brinck and Dahl (1 ♀ MZLU); São Miguel,Atalhada, [37.74, -25.58], natural forest, 4.IX.1999, leg. H. Enghoff (2 ♀♀ NHMD). Mൺൽൾංඋൺ: Campanario, Fontes, [32.69, -17.01], 1200-1400m, 16.IX.2014, leg. Kleeberg (1 J cSch); Funchal, [32.67, -16.91], 0-500 m, II.1966, leg. Palm (2 JJ 3♀♀ MZLU); Pico do Castelo, [33.08, -16.33], 17.19. V.1977, leg.Palm (2 JJ MZLU); Terreiro da Luta, [32.68, -16.91], 850 m, stream, 20.IV.1957, leg. Brinck and Dahl (1 J MZLU). SPAIN: Alicante, Orihuela, [38.08, -0.94], 20.IV.1997, leg. Lauffer (2 MNCN); Altea, Rio Algar, Alicante, [38.61, -0.05], 27.II.1999, leg. Wrase (1 J cSch); Aranjuez, Madrid, [40.02, -3.59], 520 m 3.-4.V.1988, leg. Feller (3 ♀♀ cSch); Arroyo de Jurnadiel, S Brozas, Cáceres, [39.56, -6.81], 325 m, 20.VI.1991, leg. Wrase (1 J cSch); Ávila, Avila, [40.65, -4.68], leg. Sanz (3 MNCN); Badajoz: Aljucen, [39.04, -6.33], leg. Pacheco (1 MNCN); Boadilla del Monte, NW Madrid, [40.41, -3.88], 6.VII.1996, leg. D.Wrase (1J cSch); Cabo de Gata, SE Almeria, 36.7908, -2.2513, coastal area, 5.IV.2012, leg. T. Wolsch (1 ♀ cSch); Caboalles [de Abajo], [42.95, -6.37], leg. Paganetti (1 ♀ SDEI); Cádiz, [36.53, -6.22], 16.I.1894, leg. Navarro (2 MNCN); Cádiz, Algeciras, [36.16, -5.43], leg. Arias (1 MNCN); Cádiz, Vega del Barbate, Vejer de la Frontera, [36.25, -5.95], leg. De Quirós (1 MNCN); Cornudilla, [42.68, -3.42], 13.IX.1962, leg. J.T. Skovgaard (1 J 1 ♀ NHMD); Cuenca, Uclés, [39.98, -2.86] (2 MNCN); Guipúzcoa, Arechavaleta, [43.03, -2.51], leg.Antoniol (1 MNCN); Guipuzcoa, Irun, Jaitzubia, [43.34, -1.82], 10.V.2006,leg.Anichtchenko (1J 1♀ NHMD); Huelva, Cala, [37.97, -6.31], leg. C. Bolivar (1 MNCN); Jáen Mengibar, Rio Guadalquivir, [37.98, -3.79], 330m, 16.VI.1991, leg. Wrase (1 J cSch); La Coruña, Coruña, [43.36, -8.4], leg. Ricol (2 MNCN);La Coruña, Villa Rutis, [43.28, -8.38], VII.1908, leg. Bolivar (2 MNCN); Laguna de Ruiz Sánchez, N Osuna, Sevilla, [37.41, -5.09], 26.V.1991, leg.Wrase (1 J cSch); Los Guajares, Andalusien, 36.8174, -3.5546, quarry, 6.IV.2012, leg. T. Wolsch (1 J cSch); Madrid, Escorial [40.57, -4.11] (1 MNCN); Madrid, Torrelaguna. [40.82, -3.53], leg. F. Beltran (1 MNCN); Madrid, Viñuelas [40.79, -3.34] (1 MNCN); Maranchon, Guadalajara, [41.04, -2.20], 24.XI.2005, leg.Anichchenko (1 J NHMD); Murcia, Cartagena, [37.62, -0.99], leg. S. Gomez (1 MNCN); Palencia, [42.01, -4.52], leg. Paganetti (1 J SDEI); Palmones, Cádiz, [36.17, -5.43], 29.V.1991, leg. Wrase (1 J cSch); Quero, Toledo, [39.51, -3.24], 8.III.1999, saline, leg. D.W. Wrase (1 J cSch); Serrania de Ronda, Málaga, [36.74, -5.17], 11.VI.1991, leg. Wrase (1 J cSch); Sitges, [41.23, 1.84], 22.X.1922 leg. W.Liebmann (1 ♀ SDEI); Toledo, Nahalhermosa, [39.63, -4.47], 27.VII.2007 (1 J NHMD); Tres Cantos, [40.60, -3.70], 29.-30.II.2000, leg. Anichtchenko (1 ♀ NHMD). Mൺඅඅඈඋർൺ: Palma, [39.56, 2.65], IV.1957, leg. T. Palm (1 ♀ MZLU); Rabassa, [39.54, 2.69], leg. Briet (3 JJ 1♀ NMW); Valldecousa, [39.71, 2.62], 15.-20.IV.1957, leg.T. Palm (1 J 3 ♀♀ MZLU). Cൺඇൺඋඒ Iඌඅൺඇൽඌ: Gඋൺඇ Cൺඇൺඋංൺ: Las Lagunetas, [28.01, -15.57], 22.XI.1988, leg. G. Gillerfors (1 ♀ MZLU). Tൾඇൾඋංൿൾ: Anaga Mountains, nr.Cruz del Carmen, [28.53, -16.27], 14.VI.1992, leg. Hieke & Wendt (1 ♀ ZMHB); Cumbre Dorsal Esperanza-Forest, [28.38, -16.44], 1250m, 13.IV.1992, leg. Zerche (1 ♀ SDEI); Erjos env., [28.32, -16.81], 1100 m, 7.XII.2004, leg. Skoupý (1 ♀ cSch); Genovés, [28.36, -16.74], 400m, 9.VI.1989, leg. Felies (1 J cSch); La Esperanza, [28.44, -16.37], 1200m, 18.VI.1992, leg. Hieke & Wendt (2 JJ ZMHB); Orotava, valley Aguamansa, [28.36, -16.49], 1115m, 25.XII.1995, leg. Stüben & Bahr (1 ♀ SDEI); Nr. Orotava, [28.35, -16.51], 1400m, 19.VI.1992, leg. Hieke & Wendt (2 JJ 1 ♀ ZMHB); Puerto de la Cruz env., [28.36, -16.52], 300m, 20.II.-5.III.2004, leg. E. Hajdaj (1 J cSch); Teno Alto, San Jeronimo, [28.34, -16.87], 800-900 m, 20.I.1999, leg. G. Zappi (2 JJ 1 ♀ cBor). UNITED KINGDOM: EඇGඅൺඇൽ: Chichester Harbour, [50.79, -0.96], 6.V.1947, leg. H. Last (2 JJ NMW).
Redescription. Measurements JJ (n = 6): HW = 1.62– 1.76 (1.69); HL = 1.47–1.49 (1.48); HL/HW 0.86–0.92 (0.88); PW = 2.22–2.29 (2.25); PL = 2.07–2.22 (2.16); PL/PW 0.93–0.97 (0.96); EW = 2.27–2.49 (2.37); EL = 2.20–2.33 (2.26); EL/EW 0.92–1.00 (0.96); EL/PL 1.01– 1.09 (1.05); PW/HW 1.49–1.56 (1.52); forebody length 5.76–6.02 (5.90). ♀♀ (n = 4): HW = 1.58–1.73 (1.66); HL = 1.38–1.53 (1.46); HL/HW 0.87–0.89 (0.88); PW = 1.96–2.27 (2.16); PL = 1.91–2.11 (2.05); PL/PW 0.93–0.98 (0.95); EW = 2.22–2.51 (2.42); EL = 2.11–2.42 (2.29); EL/EW 0.91–0.96 (0.95); EL/PL 1.08–1.15 (1.12); PW/ HW 1.42–1.52 (1.48); forebody length 5.40–6.07 (5.80).
Medium to large sized species; body black to brown ( Figs 9A,B View Fig ).
Head black, distinctly transverse, eyes medium size (EyL/TL: 1.88–2.06 (1.94)), microsculpture of transverse waves, no interocular punctures between anterior frontal punctures; antennae pale.
Thorax: pronotum dark brown to brown, slightly wider than long, clearly wider than head, with microsculpture of transverse waves, three punctures in dorsal row, sublateral row of pronotum with a cluster of one to three punctures situated slightly anterior of the anterior most puncture of the dorsal row; scutellum with fine setiferous punctures and with wavy to meshed microsculpture; elytra usually light brown to brown, occasionally black with paler suture and posterior margin, rarely completely black, relatively long, uniformly pubescent, quadrate or slightly wider than long, slightly longer than pronotum; legs pale, except inner face of tibia darkened.
Abdomen black to dark brown and slightly iridescent, tergites uniformly punctured, with slight iridescence.
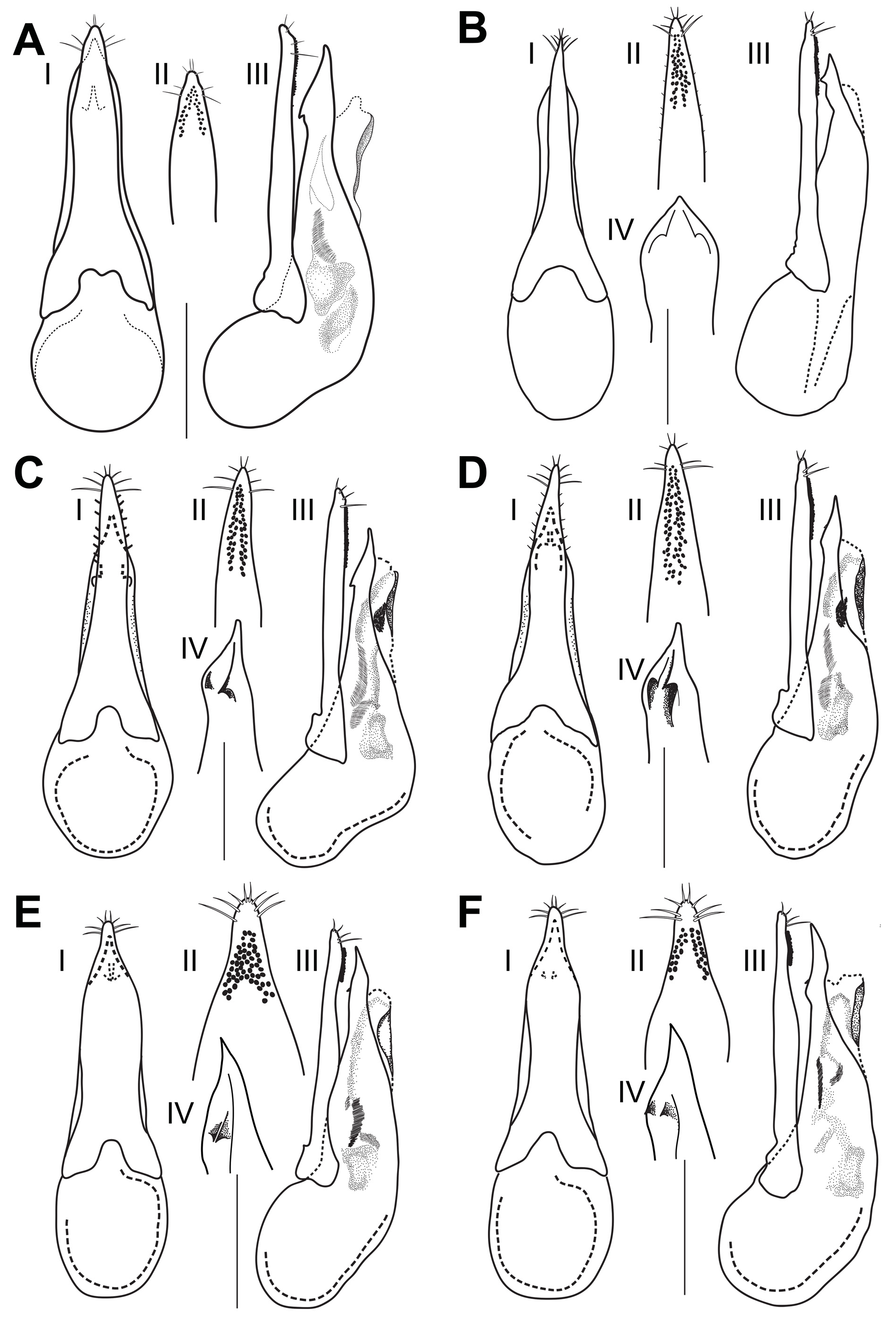
Male. Aedeagus ( Fig. 14C View Fig ): paramere elongate, narrow, lanceolate with narrowly slightly asymmetrical apical portion exceeding apex of median lobe, with numerous sensory peg setae forming two lateral longitudinal groups connected subapically; median lobe with sides of apex folded into distinct tooth situated at proximal part of fold; internal sac with a pair of very long medial sclerites, and with a pair of globular sclerites.
Differential diagnosis. Quedius hispanicus can be distinguished from all species of the Quedius s. str. except Q. pallipes , by a cluster of 1–3 additional setiferous punctures in the posterior part of the sublateral row of the pronotum. They are very rarely missing on both sides of the pronotum in Q. hispanicus . It can be distinguished from Q. pallipes only by male genitalia characters: by a slightly narrower and, in medial section, more attenuate paramere; and by a median lobe with sides of apex each folded into distinct tooth, tooth situated at base of the fold.
Synonymic notes on Q. pallipes , Q. hispanicus , Q. simplicifrons , Q. cobosi , and Q. levasseuri . Lඎർൺඌ (1849) described Quedius pallipes from the coastal town of Philippeville [now Skikda], Algeria. Fൺංඋආൺංඋൾ (1862) described Quedius simplicifrons as a very similar species from the French island of Corse. BൾඋඇHൺඎൾඋ (1898) described Quedius hispanicus , a species very similar to these, from the Spanish coastal towns of Cartagena and Cadiz. These three species have been mixed up, probably due to their very similar habitus, rendering their individual distributions unclear. Cඈංൿൿൺංඍ (1961) stated that Q. pallipes was restricted to North Africa and hypothesized that Q. simplicifrons – with Q. hispanicus as a junior synonym – could be a European subspecies of Q. pallipes without formally downgrading Q. simplicifrons to subspecific rank. Subsequently, some authors (Hൺඇඌൾඇ 1962, LඈHඌൾ 1964, Hඈඋංඈඇ 1965, Sආൾඍൺඇൺ 1970, Bඈඋ-ൽඈඇං 1976, WൾඅർH 1993) have treated Q. hispanicus as a valid species for Western European specimens, while others continued to consider it a synonym of Q. simplicifrons (Pඈඉൾ 1977, Cඈංൿൿൺංඍ 1978, Cංർൾඋඈඇං & Zൺඇൾඍඍං 1995, Sඈඅඈൽඈඏඇංĸඈඏ 2012). KඈඋGൾ (1962b) mentioned specimens from Southern Italy and North Africa as conspecific with Q. pallipes . He also reported that these were collected at higher altitudes (up to just above 1000 m) very far from the coast. Cඈංൿൿൺංඍ (1964) further described two new species based mainly on genitalia differences of the males, Quedius cobosi Coiffait, 1964 from Seron in Spain, and Quedius levasseuri Coiffait, 1964 from Corse, both similar to Q. pallipes and Q. simplicifrons / Q. hispanicus . Quedius cobosi was characterised by the presence of numerous small setae on the edge of the paramere, while Q. levasseuri was described as having paler sutural and elytral posterior margins as well as a differently shaped aedeagus, compared to Q. pallipes and Q. simplicifrons . This history implies several species- -group taxa of uncertain status, distribution and habitat preference that are similar to Q. pallipes and co-occur in the West Mediterranean. Studying a broad sample of specimens from that area, we found only two morphotypes, which differ only in the structure of the aedeagus. The first is found along the European Atlantic Coast, including Southern UK, Canaries, Madeira, Azores and inland in southern France and on the Iberian Peninsula ( Figs 14C View Fig , 19 View Fig ). These areas include the type localities of Q. hispanicus and Q. cobosi ( Fig. 19 View Fig ). The second is found in mainland Italy, on Corse and Sardinia, and in North Africa at both coastal and higher elevation sites ( Figs 14D View Fig , 19 View Fig ). These areas include the type localities for Q. pallipes , Q. simplicifrons , and Q. levasseuri . There is no transition between the aedeagi of both types. The steep cliff shoreline between Nice, France and La Spezia, Italy may be unsuitable for these species and may represent a barrier for their dispersal. As the morphological difference between both types of genitalia is clear, and in agreement with the type localities, we here propose to use Q. hispanicus for the Iberian and Atlantic coast species (type 1) and Q. pallipes for the Italian and North African species (type 2) ( Figs 14 View Fig , 19 View Fig ). Quedius cobosi syn. nov. is synonymized with Q. hispanicus .
However, barcodes from specimens across the entire distribution of both types 1 and 2 show very little variation, with no clear clustering corresponding to both genitalia types ( Fig. 3 View Fig ). A rather uniform genetic barcode among specimens morphologically corresponding to both Q. hispanicus and Q. pallipes (only 0.5% divergence, Table 2 View Table 2 ) suggests that there may be some level of mitochondrial introgression between these species. External factors such as past or present infections by endosymbionts such as Wolbachia (e.g. KඈඇൽൺඇൽൺඋൺආൺංൺH et al. 2013) cannot be ruled out as an explanation either. This should be explored more thoroughly for example with the use of nuclear genes or genomics.
The original descriptions and distribution records of Quedius hammianus Sharp, 1911 and Quedius secundus Last, 1952 , both described from the UK, match the morphology and the ecological association with coastal sites of Q. hispanicus . Quedius rufulus Blümml, 1898 , described from Granada in Spain as having darker base of the antennae, most likely represents a variant of Q. hispanicus . All three species were hitherto in synonymy with Q. simplicifrons . Under the revised concept of the Q. pallipes -group, we synonymize them with Q. hispanicus .
Bionomics. Quedius hispanicus mainly occurs in saline habitats along the Atlantic coast, such as salt marshes and the banks of estuaries in flood debris. Further south in the Iberian Peninsula and southern France the species seems to be present inland in non-saline habitats. Inland it occurs in the debris along the banks of creeks and rivers. For example, AKH has collected it in the debris near streams and rivers in the Extremadura region ( Fig. 2D View Fig ) and also in the flood debris on a salt marsh near the coast in southern Denmark. Most records are from low elevations, except a few on Madeira and the Canary Islands, where Q. hispanicus is found up to 1400 m.
Distribution. Quedius hispanicus occurs along theAtlantic coast from southern Denmark to the Iberian Peninsula where it continues to the French Mediterranean coast, with one unverified report from the northwestern Italian coast ( Fig. 19 View Fig ). In the Iberian Peninsula it occurs inland along most rivers and creeks. It also occurs on the Azores (Faial), Madeira (Madeira and Porto Santo), the Canaries (Tenerife and Gran Canaries) and Mallorca.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Quedius hispanicus Bernhauer, 1898
| Hansen, Aslak Kappel, Brunke, Adam, Simonsen, Thomas & Solodovnikov, Alexey 2022 |
Quedius cobosi Coiffait, 1964: 293
| COIFFAIT H. 1964: 293 |
Quedius hammianus
| SHARP D. 1911: 57 |
Quedius hispanicus
| BERNHAUER M. 1898: 341 |
Quedius rufulus Blümml, 1898: 216
| BLUMML E. K. 1898: 216 |