
Andinobates geminisae, Batista & Jaramillo & Ponce & Crawford, 2014
|
publication ID |
https://doi.org/10.11646/zootaxa.3866.3.2 |
|
publication LSID |
lsid:zoobank.org:pub:948CB744-768B-48AA-BF18-CB01D45A696E |
|
DOI |
https://doi.org/10.5281/zenodo.5494807 |
|
persistent identifier |
https://treatment.plazi.org/id/039187E1-FF9F-FFF1-CADD-FF5FFDC54732 |
|
treatment provided by |
Felipe |
|
scientific name |
Andinobates geminisae |
| status |
sp. nov. |
Andinobates geminisae View in CoL sp. nov.
Fig. 2 View FIGURE 2
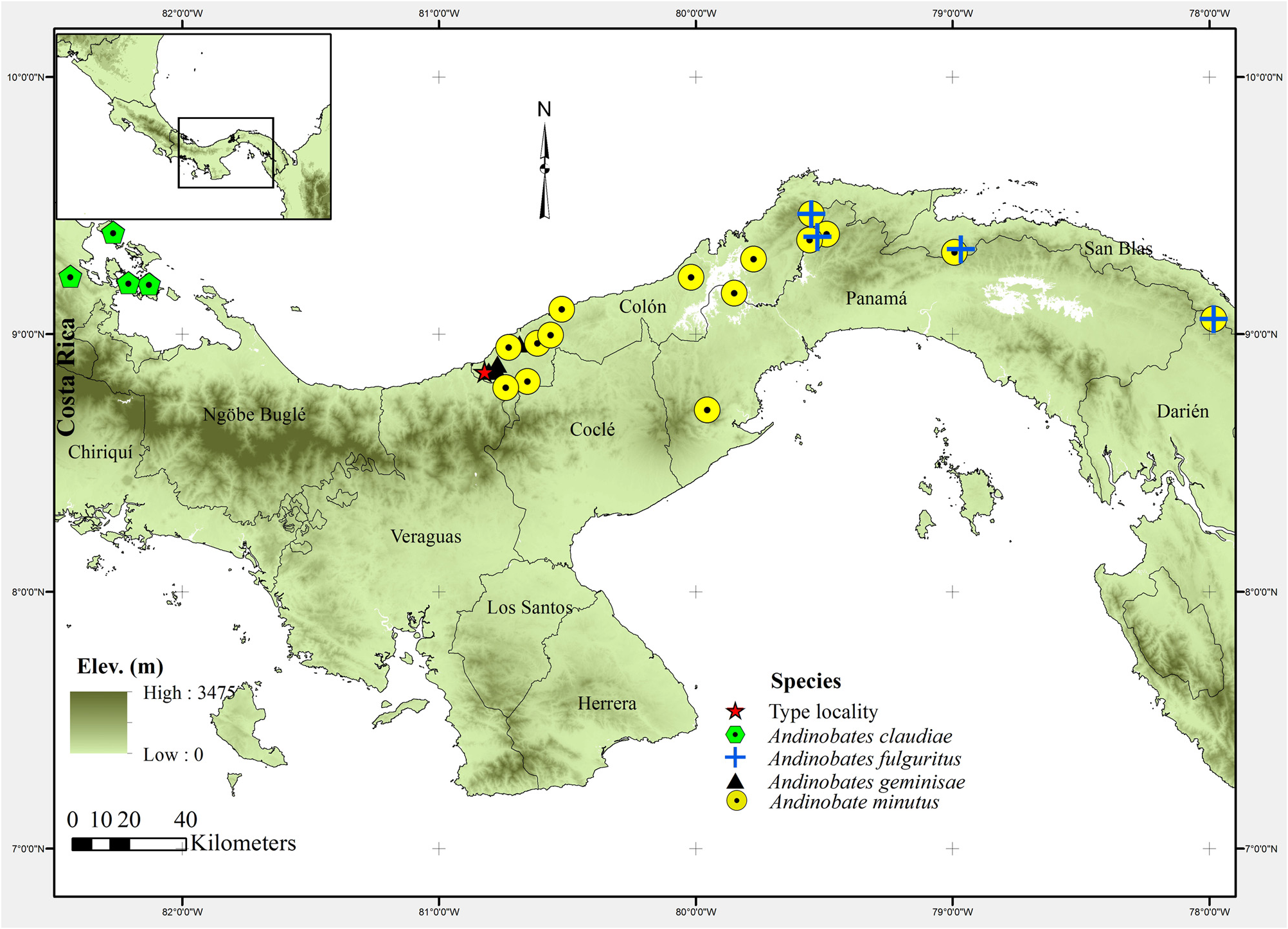
Holotype. MVUP 2428 View Materials (CH 9386: original field number of Círculo Herpetológico de Panamá), an adult male ( Fig. 2 A–B View FIGURE 2 ) collected in the headwaters of the Río Caño, Coclé del Norte , Distrito de Donoso , Colón Province, Panama, on February 21, 2011 at 09:36 AM at GPS coordinates 8.8536° N, 80.8214° W and 89 m elev. ( Fig. 1 View FIGURE 1 ) Voucher collected by Samuel Valdés and Carlos De La Cruz. GoogleMaps
Paratypes. All GoogleMaps specimens were collected between the Río GoogleMaps Coclé del Norte GoogleMaps and the Río Belén GoogleMaps basin (http:// www.hidromet.com.pa/cuencas.php; retrieved March 4, 2013), Coclé del Norte GoogleMaps , Distrito de Donoso GoogleMaps , Colón Province, Panama, and found in the leaf litter. Specimens MHCH 2073 – MHCH 2076 were collected by M. Ponce at the Manakin trail, headwaters of the Río Caño , Coclé del Norte , Distrito de Donoso , Colón Province, GoogleMaps Panama ( 8.8546° N, 80.8061° W; 82 m elev.) 7.5 km east from the Río Belén outfall on March 16, 2010. Specimens MHCH 1570 – MHCH 1576 were collected from March 15 to 16, 2010 by A. Batista, at the headwaters of the Río Chiquero , Coclé del Norte, GoogleMaps Distrito de Donoso , Colón Province, Panama ( 8.8567° N, 80.7752° W; from 100 to 136 m elev.). MHCH 2037 was collected 07 July 2009 by M. Ponce and A. Batista, at Palmilla ( Fig. 3 View FIGURE 3 ), headwaters or the Río Chiquero, Coclé del Norte, Distrito de Donoso, GoogleMaps Colón Province, Panama ( 8.8826° N, 80.7714° W; 50 m elev.) GoogleMaps .
Etymology. The specific epithet of this beautiful new species is dedicated to Geminis Vargas, the beloved wife of Marcos Ponce, for her unconditional support of his studies of Panamanian herpetology.
Diagnosis. Andinobates geminisae is characterized by the following combination of characters: (1) Tiny size, adults SVL 11.63–13.63 ( Table 2 View TABLE 2 ); (2) Dorsal coloration conspicuous; (3) dorsum and venter uniformly orange, without stripes; (4) limbs and venter brightly colored, same as dorsum; (5) Dorsal skin smooth; (6) finger I reduced and shorter than finger II; (7) finger discs III–IV moderately expanded, disc of finger III 1.11–2.38 times wider than finger width; (8) thenar tubercle slightly elongate and low; (9) toe discs II–IV weakly expanded, disc on toe III 1.03–1.71 times wider than finger width; (10) no fringes or webbing on fingers or toes; (11) outer metatarsal tubercle small and slightly pointed, inner metatarsal tubercle rounded. (12), maxillary and premaxillary teeth absent; (13) tympanum oval, posterodorsal margin hidden.
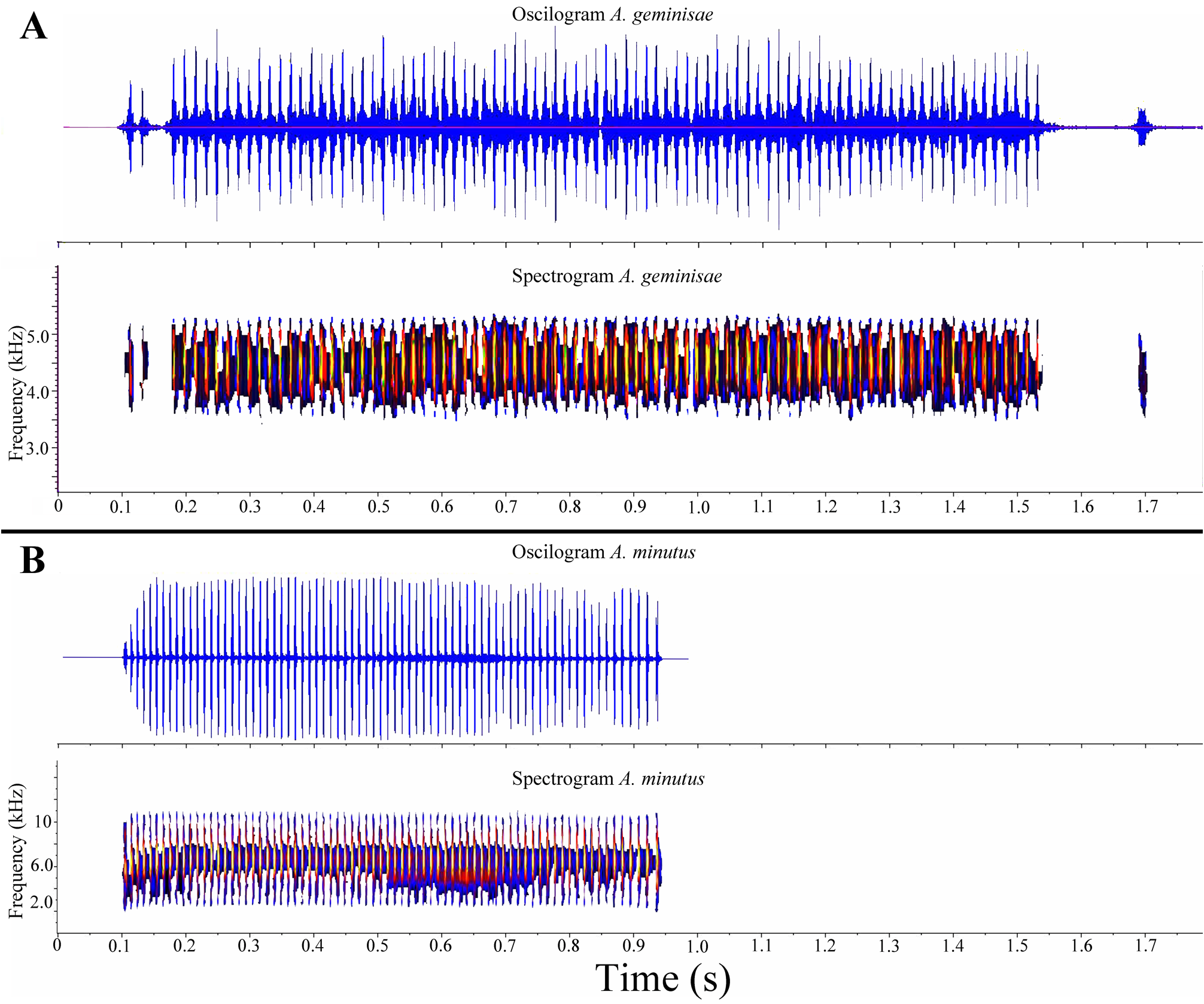
Similar species. Andinobates geminisae sp. nov. can be distinguished from other species of the genus (see table 3; Fig. 5 View FIGURE 5 ) by its predominantly uniform orange coloration through its body, smooth dorsum, and a distinctive male advertisement call (see below: vocalization description). Andinobates geminisae is similar in appearance to A. opisthomelas , but A. opisthomelas has granular skin texture (smooth in A. geminisae ), limbs, throat and belly blackish (uniform orange in A. geminisae ). Minyobates steyermarki ( Rivero 1971) also has reddish coloration, but has the first finger as long as the second or greater ( A. geminisae finger I shorter than finger II). Andinobates minutus is closely related to A. geminisae ( Fig. 5 View FIGURE 5 ), which reaches the western limit of the distribution of A. minutus , but despite their very close geographical distribution ( Fig. 1 View FIGURE 1 , 6 View FIGURE 6 ), the coloration of A. geminisae contrasts with that of A. minutus (see Fig. 2 View FIGURE 2 , 5B View FIGURE 5 ). Furthermore, Andinobates minutus , A. claudiae and A. fulguritus have higher dominant frequency in their advertisement calls than A. geminisae ( 4.8–6.4 in the three first species, versus 4.4–4.5 kHz in A. geminisae ; see Brown et al. 2011; Fig. 7 View FIGURE 7 ); Andinobates minutus , A. claudiae and A. fulguritus also have fewer pulses per note than A. geminisae ( 28–71 in the three first species, versus 87–88 in A. geminisae ; see Brown et al. 2011). Oophaga pumilio and O. vicentei are larger than A. geminisae ( 11.63–13.63 mm SVL); O. pumilio is usually> 17 mm SVL ( Batista & Köhler 2008; Köhler 2011) and O. vicentei 19–21 mm SVL ( Jungfer et al. 1996, Lötters et al, 2007). The dorsal color pattern of O. pumilio is usually red with blue legs, however it present a great color variation, from green, yellow, blue or red, uniform or with dark blotches (8 C–D). Two populations of uniformly red O. pumilio are known in Panama, those from Cerro Tebata ( Fig. 8 E–F View FIGURE 8 ) and from Solarte Island. However, these populations are larger (> 17 mm SVL; Batista & Köhler 2008) than A. geminisae . Oophaga vicentei is mainly a tree-dwelling species ( A. geminisae is terrestrial). The dorsal color of O. vicentei near the type locality of A. geminisae varies from brown, reddish, yellow or blue, all with dark blotches or a reticulated pattern, not uniform, and the venter is sky blue ( Fig. 8 A–B View FIGURE 8 ). The call duration in O. pumilio is 0.04– 0.07 s (Myers & Daly 1976), and the call duration of O. vicentei is 0.14– 0.20 s (MHCH 2983, a male recorded at Donoso, Colón, Panama, 8.82246° N, - 80.70504° W, 294 m elev., 22 July 2011, 07:30, 23 °C), while the call duration of A. geminisae is much longer ( 1.60 s).
Measurements of the holotype in mm. (See Table 2 View TABLE 2 ) SVL 13.63, HW 4.19, HL 4.21, IND 1.84, IOD 1.49, ED 1.80, EYE-NAR 1.03, TYMP 0.77, TL 5.86, FOOT 5.08, FAL 3.32, HaL 2.86, 3FW 0.38, 3FD 0.48, 3TW 0.38, 3TD 0.45, 4TW 0.38, 4TD 0.50, BW 4.40, LIIF 1.25 LIF 0.85, IND/SVL 0.13, TL/SVL 0.43, HL/SVL 0.31, HW/SVL 0.31, HL/HW 1.00, ED/SVL 0.13, TD/SVL 0.06, SL/SVL 0.08, HAL/SVL 0.21, FL/SVL 0.37, FAL/ SVL0.24, BW/SVL 0.32, 4TD/4TW 1.32, 3TD/3TW 1.18, 3FD/3FW 1.26, LIIF / LIF 1.47.
Description of the holotype ( Fig. 2A–B View FIGURE 2 ). An adult male with slender body; smooth dorsal skin, venter coarsely areolate; eye length longer than snout; ratio SL/ED 57%; tympanum of moderate size, ratio TD/ED 43%; tympanum distinguishable, annulus tympanicus oval and partially visible through skin, posterodorsal margin hidden, positioned closely behind orbit and upper jaw; head equally wide as long (ratio HL/HW: 1.00); head width 31% of SVL; snout nearly rounded from above and rounded in profile; nares situated near tip of snout and laterally directed, visible ventrally and in front view, barely visible dorsally; internares distance 1.84 mm, 44% of head width; canthus rostralis rounded, loreal region flat; paired vocal slits present and each is located adjacent to body of the mandible between the tongue and the jaw articulation; tongue elongate, median lingual process absent; hands moderate in size, 21% of SVL; relative lengths of adpressed fingers I<II<IV<III; finger IV slightly longer than II, finger II reaching the disk on finger IV when adpressed; finger III disk 1.26 times wider than distal end of adjacent phalanx; palmar tubercle rounded, slightly elevated, and slightly larger than thenar tubercle; thenar tubercle low and elongate; subarticular tubercles rounded and globular; no supernumerary tubercles; palmar and plantar accessory tubercles absent; no nuptial pads; no fringes on fingers; hind limbs of moderate lengths, TL 43% of SVL; relative lengths of adpressed toes I<II<III<V<IV; when adpressed, tip of toe I reaches to the proximal tubercle of toe II; disc of toe IV weakly expanded, 1.32 times wider than distal end of adjacent phalanx; no fringes on toes; one to three nonprotuberant subarticular tubercles present (one on toes I and II, two on toes III and V, three on toes IV, subarticular tubercles of toe IV weakly defined); inner metatarsal tubercle elongated; outer metatarsal tubercles small rounded and conical and smaller than inner; plantar tubercles small and rounded; tarsal ridge absent; toe disks weakly expanded; hands and feet without webbing.
Variation. Eleven adult specimens collected including the holotype and one juvenile; SVL: 12.68 ± 0.87 (11.63–13.63); males: 12.70 ± 0.74 (11.63–13.63; n: 8); females: 13.25 ± 0.05 (13.20–13.29; n: 3); one juvenile: 10.80. Females slightly bigger than males .
Color in life ( Fig. 2 View FIGURE 2 ). Coloration of most specimens fit with the color description of the holotypes, as follows: dorsum uniform Chrome Orange (74), interorbital area slightly suffused with dark pigmentation; lower tympanum area, junctions of elbow and knee Fawn Color (258); fingers and toes Glaucous (272), suffused with Spectrum Orange (9); plantar and palmar areas Prout’s Brown (47); subarticular tubercles Pale Neutral Gray (296); junction between axila and body Fawn Color (258); throat and belly Spectrum Orange (9); lips of lower jaw Chestnut (30); iris black.
Color in preservative. Dorsum posterior to supraorbital region and anterior to the lumbar region Light Neutral Gray (298); front of head and limbs Russet (44); ventral surfaces Hazel (26) with the glandular spots Lavender (202). In preservative specimens MHCH 1991 and MHCH 1993 have little dark spots on throat and belly. Specimens MHCH 1991, MHCH 1571, MHCH 1573 and MHCH 1575 have dark pigmentation on throat.
Tadpole. Known only from photo ( Fig. 2C View FIGURE 2 , not collected). One individual (undetermined sex) carried on back one dark tadpole, observed on 16 March 2010 at 13:50 hrs.
Vocalization. We recorded one male on 15 March 2010 in the Distrito de Donoso GoogleMaps , Colón Province, Panama ( 8.8546° N, 80.8061° W) at 11:35 hrs with air temperature of 25.9ºC. The calls consisted of a buzz-like call similar to other members of the A. minutus species group ( Brown et al. 2011). Two calls were recorded ( Fig. 7 View FIGURE 7 ); details of the first call as follows (with information on the second call provided in parentheses): calls consisted of 87 (88) pulses, call duration of 1.60 s ( 1.52 s), with an interval between the two calls of 59.4 s, with a total call rate of approximately two calls per minute. The rise time was 5.64% (8.14%) of the total call duration. The peak frequency band ranged from 4.12 to 4.74 kHz (4.03–4.80 kHz), with the dominant frequency at 4.52 kHz (4.4 kHz). Each pulse length was 7.8 ms (7.7 ms), and the pulse interval on average was 10 ms (9.5 ms). The call began with one or two pulses spaced by a pause of 42.0 ms (52.0 ms) from subsequent pulses, after which the pulse intervals remained relatively constant at 8.40 ± 1.6 ms. Before the final pulse there was an interval of silence lasting 152.0 ms (88.0 ms) followed by a click that terminated the call.
Distribution and natural history ( Figs. 1 View FIGURE 1 , 3 View FIGURE 3 & 6 View FIGURE 6 ). Andinobates geminisae is known only from the Río Belén basin (http://www.hidromet.com.pa/cuencas.php; retrieved March 4, 2013; Fig. 1 View FIGURE 1 and 6 View FIGURE 6 ). Most A. geminisae were found in nearly pristine forest with occasional perturbations by people looking to establish settlements ( Fig. 3 View FIGURE 3 ). All individuals were found on ridges of small hills, suggesting that A. geminisae prefers upland drier areas over flat, moister low-lying areas. Most frogs were found exposed on top of the leaf litter, nearby tree roots or near the entrance of cavities under rocks or logs. The bright coloration of A. geminisae contrasts sharply with the forest floor, allowing the investigator to see the frogs easily. In an area of 40 m 2 ( 8.8567° N, 80.7752° W; 136 m. elev.) we observed approximately 15 individuals in 45 minutes. Calling males of A. geminisae were heard from 08:00 to 14:00, suggesting that this species calls actively throughout the day during the breeding season. Potential areas for reproduction used by this species may include phytotelmata found at the type locality, e.g., water-filled tree hollows plant axial water (e.g. in Heliconia spp. ), and dry leaves filled with rain water, though no tadpoles were found. Dendrobatid species sympatric with A. geminisae included Colostethus pratti , Dendrobates auratus , Oophaga vicentei , Phyllobates lugubris , and Silverstoneia flotator .
Conservation status. Despite the fact that most of the terrestrial habitat in the Donoso region is comprised of primary forest, during recent decades the area has been impacted by deforestation for pasture and agriculture land by non-indigenous colonists (see Fig. 3 View FIGURE 3 ), as well as migration of indigenous peoples from the Comarca Ngöbe Buglé to establish new settlements. More recently, multinational mining companies have begun surveying the region with intentions to build a copper mine. Therefore, the development and implementation of conservation action plans should be a high priority to assure the survival of A. geminisae .
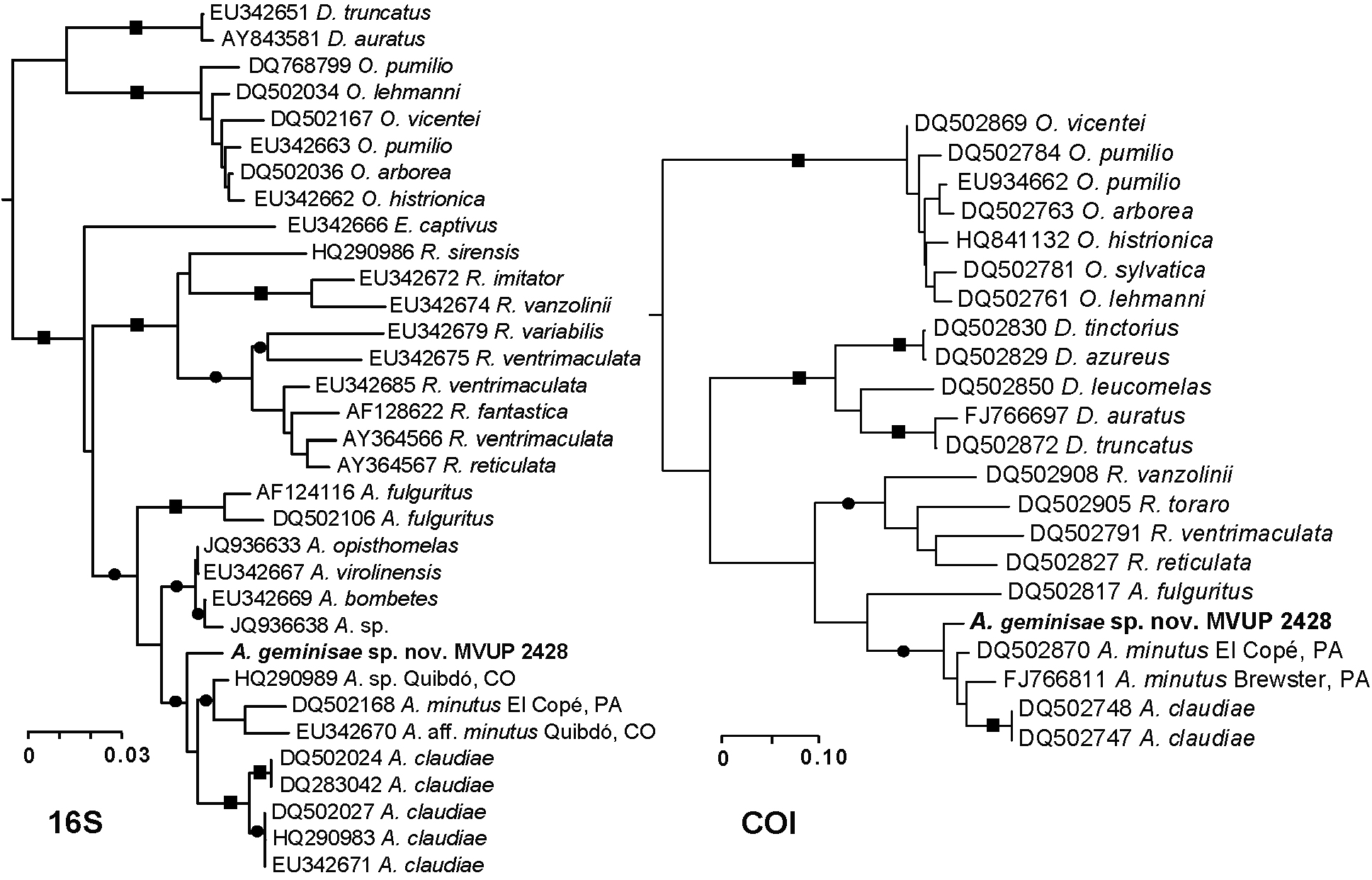
Genetic distance results ( Tables 4–5, Fig. 4 View FIGURE 4 ). Assuming HKY distances at the COI marker, A. geminisae (MVUP 2428) showed 5.6% divergence (Table 4) from A. minutus ( DQ502870 View Materials , KRL 0790, Parque Nacional General de División Omar Torrijos Herrera, Coclé Province, Panama), 8.5% from a second A. minutus sample ( FJ766811 View Materials , CH 9887, Cerro Brewster, Distrito de Panamá, Panama Province, Panama), 8.6% divergence from A. claudiae ( DQ502747 View Materials , USNM-FS 59979, Isla Popa, Bocas del Toro Province, Panama) and 18.7% from A. fulguritus ( DQ502817 View Materials , MHNUC 340, Bahía Solano, Chocó, Colombia). (Adding a G-rate heterogeneity parameter with values estimated by ML, these distances would be 8.4%, 15.5%, 15.9% and 73.5%, respectively.) For the 16S data, A. geminisae (MVUP 2428) was most similar to A. sp. ( HQ290989 View Materials , TNHC FS 4943, Quibdó, Chocó, Colombia) and A. minutus ( DQ502168 View Materials , KRL 0790, Parque Nacional General de División Omar Torrijos Herrera, Coclé Province, Panama) with 2.5% HKY distances from either sample. Relative to other Colombian congeners, the new species showed 2.7% divergence from the two genetically identical A. opisthomelas ( JQ936633 View Materials ) and A. virolinensis samples ( EU342667 View Materials ), and 3.1% divergence to A. bombetes ( EU342669 View Materials ). At 16S, A. geminisae showed 3.4% divergence from A. claudiae ( DQ283042 View Materials and DQ502027 View Materials ), and 3.6% divergence from Colombian samples A. cassidyhornae ( JQ936638 View Materials , GECOH 1534S, Antioquia, Colombia) and A. aff. minutus ( EU342670 View Materials , TNHC FS 4943, Quibdó, Chocó, Colombia). Finally, A. fulguritus showed 4.9% ( AF124116 View Materials ) to 5.5% ( DQ502106 View Materials ) divergence from the new species.





| AM |
Australian Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |