
Therochaeta caudata ( Rioja , 1963 )
|
publication ID |
https://doi.org/10.5252/z2013n2a7 |
|
DOI |
https://doi.org/10.5281/zenodo.7191288 |
|
persistent identifier |
https://treatment.plazi.org/id/03985007-F25A-FFEB-02F0-FD1EFB834D2B |
|
treatment provided by |
Felipe |
|
scientific name |
Therochaeta caudata ( Rioja , 1963 ) |
| status |
|
Therochaeta caudata ( Rioja, 1963) n. comb.
( Fig. 2 View FIG )
Yliphagus caudatus Rioja, 1963: 191-195 , figs 94, 95.
Flabelliderma caudata – Salazar-Vallejo 1981: 112, 113, fig. 12. — Hendrickx et al. 1997: 22, 23.
Ilyphagus caudatus View in CoL – Salazar-Vallejo 1989: 52.
MATERIAL EXAMINED. — Eastern Tropical Pacific.– Four specimens, two ( UNAM 4524 ) and two ( UNAM 4549 ), Mazatlán Bay, Sinaloa, México ( 23°10’N, 106°24’W), 17 m, 8.XII.1996 GoogleMaps . Three specimens, two ( UANL) and one specimen ( ECOSUR), Estero de Urias ( 23°12’N, 106°23’W), Mazatlan, Sinaloa, Mexico, intertidal, 15.I.1979, A. Rutgers coll. GoogleMaps
DISTRIBUTION. — This is a shallow water species that has been collected in 0-24 m in the southwestern coast of the Baja California peninsula and in Mazatlan, Sinaloa.
DESCRIPTION
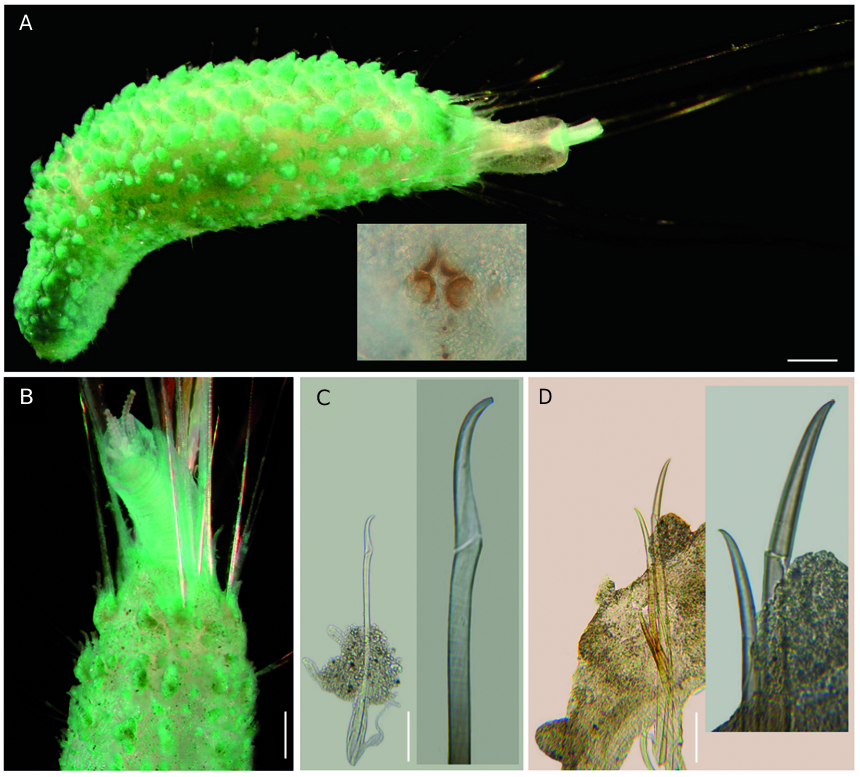
Largest specimen (UNAM 4524) cylindrical,globose, tapered posteriorly ( Fig. 2A View FIG ). Body cross-section oval, slightly compressed; 8 mm long, 1 mm wide, cephalic cage 4 mm long, 21 chaetigers.Tunic thin with fine sediment particles. Body papillae filiform capitate, with large amount of sediment towards its base forming tubercles of different sizes, larger dorsally, about three-four rows per segment.
Cephalic hood exposed, narrow, made by two rings ( 2 mm long); basal ring rugose, finely annulated, at least five times longer than distal ring; distal ring transparent, smooth. Cephalic hood margin crenulated, with small papillae.
Anterior end observed by dissection in another specimen (UNAM 4549). Prostomium low cone with four dark brown eyes ( Fig. 2A View FIG ); two inner pigmented spots placed laterally to the anterior eyes. Caruncle not detected. Palps thick, corrugated; palp keels not seen.
Branchiae cirriform, arranged into two marginal dorsolateral rows, one on each side, about 20 filaments with ciliary girdles ( Fig. 2B View FIG ). Palps and branchiae apparently of the same size when contracted.
Cephalic cage chaetae half as long as body length, four times longer than body width. Cephalic cage made only by chaetigers 1-2, with chaetiger 2 slightly displaced dorsally.Chaetiger 3 with chaetae long but less than half those present in chaetiger 2. Cephalic cage chaetal arranged in short, lateral series. Four to five chaetae per bundle.
Anterior dorsal margin of first chaetiger papillated; papillae sessile, not placed over any projection. Chaetigers 1-2 with long papillae over chaetal lobes, longer than following chaetigers. Post-cephalic cage chaetigers not markedly elongated; only chaetiger 3 longer. No constriction behind those chaetigers forming the cephalic cage; parapodia of chaetigers 1-2 better developed than posterior ones. A dorsal, crescentic glandular pad over chaetiger 2, slightly extended forward over chaetiger 1. Chaetal transition from cephalic cage to body chaetae abrupt; pseudocompound neurohooks from chaetiger 4. Gonopodial lobes not seen.
Parapodia better developed in chaetigers 1-2; following parapodia poorly developed; chaetae emerging from body wall. Notopodial papillae longer than neuropodial ones. Noto- and neuropodia close to each other.
Median notochaetae arranged as lateral tufts. All notochaetae capillary multiarticulated, brittle, as long as about 1/4 body width in middle segments, articles long throughout the chaetae. Median neuropodia ventrolateral. Neurochaetae multiarticulated capillaries in chaetigers 1-3, one-two pseudocompound hooks in chaetigers 4-8 ( Fig. 2C View FIG ), from chaetiger 9 falcate, anchylosed neurohooks, mostly two per bundle ( Fig. 2D View FIG ), to the end of body.
Posterior end thick, conical; anus terminal without anal cirri.
REMARKS
Therochaeta caudata n. comb. and T. fauchaldi n. sp. are the only species in the genus without an anterior shield. They differ in the relative development of the sediment tubercles; in T. caudata n. comb. there are fewer per segment with the dorsal ones larger, whereas in T. fauchaldi n. sp., they are more abundant and of similar size.
Rioja (1963: 191) described Ilyphagus caudatus Rioja, 1963 from off Western Baja California Sur, Mexico, with specimens 10-12 mm long, 3-4 mm wide, collected in 21 m, but his type materials were lost ( Salazar-Vallejo 1989). The finding of some intertidal materials collected in Mazatlán, Sinaloa, which were regarded as belonging to the same species, allowed for the transfer of the species to Flabelliderma ( Salazar-Vallejo 1981: 112, 113); because of the presence of pseudocompound neurohooks in anterior chaetigers it keyed out in that genus. This transfer was incorrect as the species belonging to Flabelliderma Hartman, 1969 have multiarticulate neurohooks from chaetiger 2 to the end of the body, although those chaetae have been regarded as pseudocompound, they are actually multiarticulate with a distal piece or crest ( Salazar-Vallejo 2007). The original combination was later retained; Rioja’s species has been regarded as a member of Ilyphagus ( Salazar-Vallejo 1989: 52) but that placement is also questionable due to the different development of papillae because they form large fine sediment tubercles in Rioja’s species, whereas in Ilyphagus Chamberlin, 1919 they do not form tubercles ( Salazar-Vallejo 2012a). Besides, Rioja’s materials were from shallow water and Ilyphagus is a deep-water genus.
On the basis of body shape, papillae, and chaetal features, I. caudatus may fall within Bradabyssa Hartman, 1967 . However, the species belonging in Bradabyssa lack a well-developed cephalic hood, and its anterior end is barely exposed. The illustrations by Rioja indicate a very long, thin cephalic hood; together with papillae and chaetal features which indicate its affinities with other Therochaeta species. Thus, the transitional pseudocompound neurohooks were likely overlooked in the original description; taking the other features into consideration, I. caudatus must be transferred to Therochaeta , and hence the new combination. However, because these specimens are not from the type locality, they cannot be designated as a neotype; this must wait until material becomes available.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Therochaeta caudata ( Rioja , 1963 )
| Salazar-Vallejo, Sergio I. 2013 |
Flabelliderma caudata
| HENDRICKX M. E. & PEREZ M. C. & BARRAGAN J. & UBACH M. N. 1997: 22 |
| SALAZAR- VALLEJO S. I. 1981: 112 |
Yliphagus caudatus
| RIOJA E. 1963: 191 |