
Sabellaria guamare, Santos, André Souza Dos, Brasil, Ana Claudia Dos Santos & Christoffersen, Martin Lindsey, 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3881.2.2 |
|
publication LSID |
lsid:zoobank.org:pub:CE5F4D90-4AC0-44F1-8C29-0BC5E92043C8 |
|
DOI |
https://doi.org/10.5281/zenodo.5625856 |
|
persistent identifier |
https://treatment.plazi.org/id/039C87DF-7E21-C938-FF78-FE5DFE9BF878 |
|
treatment provided by |
Plazi |
|
scientific name |
Sabellaria guamare |
| status |
sp. nov. |
Sabellaria guamare sp. n.
Figures 5 View FIGURE 5 A–F, 6A–F, 7A–M and 8A–D, Tables 1 View TABLE 1 and 2 View TABLE 2 .
Material examined. Total: 22specimens. Type series: State of Rio Grande do Norte, Guamaré City: Projects Caracterização Ambiental da Bacia Potiguar and Monitoramento Ambiental dos Emissários Submarinos do Pólo Industrial de Guamaré . St. BPOT 24DG (05º3.561’0’’ S036º28.57’ 0’’W), 77,6 meters of depth. Holotype [CIPY- POLY 1689]: 1 complete spec. 21/05/2004, 55 mm long. Paratypes: Paratype 1, [CIPY-POLY 1690]: 1 complete spec. 21/05/ 2004 in a good state of preservation. Paratype 2 [CIPY-POLY 1691]: 1complete spec. 21/05/2004. Paratype 4 [CIPY-POLY 1692]: 1incomplete spec. 21/05/2004. Paratype 5 [ POLY 1693]: 1 incompletespec. 21/05/ 2004. Paratype 6, [CIPY-POLY 1694]: 1 incomplete spec. 21/05/2004. Paratype 7 [ POLY 1695]: 1 incomplete spec. 21/05/2004. Paratype 8 [ POLY 1696]: 21/05/2004. Paratype 9[CIPY-POLY 1697]: 21/05/2004. Paratype 10 [CIPY-POLY 1698]: 1 incomplete spec. 21/05/2004. Paratype 11 [xxx-POLY 1699]:1 incomplete spec. 21/05/ 2004. Paratype 12 [CIPY-POLY 1700]: 1incomplete spec. 21/05/2004. Paratype 13 [CIPY-POLY 1701]: 1incomplete spec. 21/05/2004. Paratype 14 [CIPY-POLY 1702]: 1incomplete spec. 21/05/2004. Paratype 15 [ POLY 1703]: 1 incompletespec. 21/05/2004. Paratype 16 [CIPY-- POLY 1704]: 1 incomplete spec. 21/05/2004. Paratype 17 [CIPY-POLY 1705]: 1 incomplete spec. 21/05/2004. Paratype 18 [CIPY-POLY 1706]: 1 incomplete spec. 21/05/ 2004. Paratype 19 St. BPOT 25DG. (05º20’0’’S 036º40’0’’W)[MN-POLY 1707]: 1 incomplete spec. 19/05/2004, 5 m. Paratype 20 St. BPOT 25DG (05º20’0’’S 036º40’0’’W). [MN-POLY 1708]: 1 incomplete spec. 19/05/2004, 5 m. Paratype 21 St. E16DG (05º3.561’0’’ S 036º29’0’’ W)[MN-POLY 1709]: 1 complete spec. 21/05/2004, 10 m, 3 mm long.
Stubs for SEM. Paratype 3 [CIPY-POLY 1710]: 1 incomplete spec. 21/05/2004. Paratype 4 [CIPY-POLY 1711]: 1 incomplete spec. 21/05/2004. Paratype 12 [CIPY-POLY 1712]: 1 incomplete spec. 21/05/2004. Paratype 13 [CIPY- POLY 1713]: 1 incomplete spec. 21/05/2004.
Diagnosis. This is a unique species of Sabellaria in possessing a prolonged form of the medium cirrus. This structure shows 3 small prolongations with extremity gauged, and small black eyespots agglomerated in some parts of the medium organ. The organization of the opercular crown is also exclusive. The opercular crown is organized into: outer paleae with small teeth (five to six along each distal margin), distal teeth present, with 2–3 distal teeth on each side of dominant tooth, progressively smaller toward outer margins on either side of dominant tooth. Middle paleae alveolate, strongly geniculate, with falciform blades lightly blunted, the inner portion positioned toward inner series of opercular crown; weakly excavate inner face of thecal bands, at lower portion; one to two large teeth on each side of dominant tooth.
Description. Holotype. Complete specimen in good conditions with 45 chaetigers, total length 55 mm, length of peduncle 3 mm, width of peduncle 3 mm, length of thorax 2.5 mm, width of thorax 2 mm, and length of cauda 3 mm ( Figures 5 View FIGURE 5 A, B; 6A, B and 7K, M). Anterior end of opercular stalk and crown with light mid-ventral indentation, divided into symmetrical halves ( Figures 5 View FIGURE 5 C and 6C). Opercular paleae yellow-gold with vitreous luster organized in rows surrounding prostomium and peristomium in three apparent concentric rows differing in shapes and length. Outer paleae forming 28–30 pairs on each side, with flattened, oblong, thecate blades; plume with three pairs of teeth on side of dominant tooth ( Figures 5 View FIGURE 5 C, D and 7A). Medial teeth denticulate. Medial plume on distal margin of blades with serrate distal teeth, with1 distal tooth on each side of dominant tooth; these progressively smaller toward outer margins on either side of dominant tooth ( Figure 8 View FIGURE 8 A, B). Blades bent slightly outward from longitudinal axis of shaft; distal lateral margins directed posteriorly; inner surface of blade with many fine closely spaced striations ( Figure 8 View FIGURE 8 C, D). Each striation consists of series of small teeth (occurring along a limited region).Twelve pairs of middle paleae on each side of the symmetrical halves. Middle paleae alveolate, strongly geniculate, with falciform blades slightly blunt, curved 90° ( Figure 7 View FIGURE 7 B), with axis towards inner series on opercular crown; weakly excavate innerface of thecal bands; with one to two divisions in apical region, and with one to two distal teeth in each side of dominant tooth and series of tiny extensions. Distal ends of paleae almost erect, tapering to distal tips, with weak excavation on inner face of thecal bands ( Figure 5 View FIGURE 5 ). Paleae of inner series strongly geniculate; proximal upper teeth along margin with thecal bands weakly excavate ( Figure 7 View FIGURE 7 C), slanting upward from horizontal plane; blade thicker and wider toward base, terminating distally in bluntly rounded point, series of distal teeth along margin, occurring along a limited region ( Figure 7 View FIGURE 7 C, D). Opercular peduncle with 3 pairs of dorsal (nuchal) spines on either side of mid-line ( Figure 7 View FIGURE 7 D). Anterior margin of peduncle with flattened big spaced palps in series attached to peristomium ( Figures 5 View FIGURE 5 A–C and 6C). Peduncular bases with a series of 6 longitudinal rows of compound feeding tentacles (12–15 in transverse rows) on ventral and dorsal margins of buccal cavity attached to peristomium ( Figure 5 View FIGURE 5 C, D). Prostomium fused with peristomium and limited to a region situated in front of the mouth ( Figure 5 View FIGURE 5 C), bearing pair of grooved prehensile tentacles with conspicuous medium cirrus, with prolonged form positioned in front of grooved buccal palps; small black eyespots agglomerated in some parts of the medium organ ( Figure 6 View FIGURE 6 , F). Peristomium lips beginning anterior to the mouth, ending at the base of the compound feeding tentacles ( Figure 5 View FIGURE 5 D). Thoracic region consists of a U-shaped building organ, and two segments with spiraled capillary chaetae ( Figure 7 View FIGURE 7 E). Thoracic segments with neuropodium only, without notopodium, and conical palps located on each side of opercular peduncle (ure 5A, B, and E). Cirri of first thoracic segment triangular, bent toward ventral side of body, quite similar to buccal cirri. Cirri of second thoracic segment triangular, elongate, bent toward dorsal side of body, on each side of the mid-ventral indentation of operculum. Parathoracic region with 3 segments and small biramous parapodia ( Figure5 View FIGURE 5 A, B, and E). Notopodium and neuropodium with shaves of chaetae with lanceolate tips (“oar-shaped”) ( Figure 7 View FIGURE 7 F, H) and thin spiraled capillary chaetae ( Figure 7 View FIGURE 7 G, I) with slightly curved tips positioned between chaetae with lanceolate tips. Abdomen with 18–20 segments and tori in numerous segments ( Figure 6 View FIGURE 6 C); dorsal digitiform branchiae in the five anterior abdominal segments, being reduced in size at segment 15 and disappearing in last segment ( Figure 5 View FIGURE 5 F) tori with chaetae positioned in a single row. Notopodial uncini bipectinate, with 6 rows of superposed teeth, neurochaetal conical cirri arising from base of bundle in first abdominal segment; cirri diminishing in size posteriorly and disappearing on posterior-most segments ( Figures 7 View FIGURE 7 J,L). Caudal region septate, composed of numerous achaetous segments. Cauda bent towards ventrum ( Figures 5 View FIGURE 5 A, B and 6C). 16–20 anal septations. Eyespots brownish to reddish present in each expansion of the filaments of compound feeding tentacles (or branchial filaments). Black pigmented eyespots in opercular peduncle along inner face of serial conic palps, extended to midventral indentation; peristomium between compound feeding tentacles, medium cirrus in base of upper edge; parathoracic region with a large stain composed of numerous eyespots in ventral side of third setiger; few eyespots in caudal region. Red glandular spots on abdominal region, on paired cirri, and on neuropodial pads of chaetigers. Red eyespots form “C”-shaped pads in first abdominal chaetigers. Eyespots increase in number and concentration beyond abdominal chaetiger 10.
Variations. The specimens examined present great variations in the number of eyespots. Some specimens do not have the brownish to reddish eyespots, or have them in lower concentrations than in other samples. The specimens found at the depths of 5–7 m are generally more pigmented than the specimens collected at 7–77 meters. These differences could represent sexual dimorphisms because we observed that specimens collected in 77 meters have a lot of black eyespots and others only a few. Lomonaco et al. (2011) observed that males of Sabellaria nanella and S. wilsoni are more pigmented than females. The number of abdominal segments does not vary incomplete animals as shown below in Table 1 View TABLE 1 .
Type locality. Brazil: Rio Grande do Norte State: Guamaré Bay.
Etymology. The species is named after Guamaré Bay, the type locality of this species.
Remarks. Sabellaria guamare n. sp. belongs to the group of the honeycombed species of Nishi et al. (2010) ( Table 2 View TABLE 2 ). This group of species of Sabellaria possesses three kinds of paleae. These species have some distal teeth that are paired in relation to the central teeth. The species of this group always have honeycomb bed medium paleae, light or deeply excavated, that could be bent outwards. The inner paleae are honeycombed or expanded anteriorly ( Figure 8 View FIGURE 8 C), extremely geniculate, and could be reflected outwards or sometimes could be curved into an angle of 45°.
Among the alveolate species, only S. bellis Hansen 1882 , S. guamare n. sp., and S. moorei (Monro 1933) , possess the outer paleae with 2–3 big teeth on each side of the dominant teeth. Only S. guamare n. sp. shows a rotated medium honeycombed palea. Therefore, the dominant teeth of S. guamare n. sp. are longer and have the distal end with small teeth of similar size. In S. moorei and S. bellis they are triangular and decreasing in size.
S. guamare sp. n. constructs solitary tubes made by mud sand on sand substrata, but some specimens were found constructing their tubes around the tubes of other polychaetes, such as Loimia View in CoL sp., S. wilsoni Lana and Gruet, 1989 View in CoL , and S. nanela Chamberlin, 1919 . S. guamare sp. n. inhabits depths from three to 77 meters. This species lives in estuarine areas, and is the only gregarious sabellariid that doesn’t use any hard substrata to construct their sand tubes. This species is known only from the type locality.
Species Nuchal Outer paleae Middle paleae Inner paleae Type locality Distribution Reference
spine
According to Kirtley (1994), the genera and species are defined based on the configuration in the opercular stalk and crown, the number and kinds of paleae of the opercular crown, differences in the geometry and superficial ornamentation of the thecae and paleae, presence of nuchal hooks or spines, presence or absence of a median organ (or aboral cirrus), form of the oral feeding tentacles, shape and relative length of the prostomial tentacles, shape and size of the building organ, shape and number of cirri, details of associated chaetal bundles on the first and second chaetigerous segments, number of parathoracic segments, and presence or absence of dorsal branchiae on some abdominal segments. This work shows that the paleae of some taxa are composed of series of small teeth, and Kirtley named these structures tiny extensions ( Figure 8 View FIGURE 8 A–D). These tiny extensions should be observed for other sabellariids as well. The major illustration provided in the revision proposed by Kirtley (1994) and subsequent articles shows a majority of paleae with structures described as lines, tiny extensions or striations.
Our results show that the paleae of all species of Sabellaria View in CoL examined in this study, as well as species described by Nishi et al. (2010) and Santos et al. (2011), always have a series of teeth and the palea of each species has its own form. Consequently, if Kirtley (1994) and other authors had examined and compared the paleae of different taxa with SEM, they could have concluded that the striations are in fact a series of teeth and could be a new character used in the taxonomy of Sabellaria View in CoL .
The major abundance and diversity of species was concentrated around the depth of 77.5 meters. At this depth more than 25 samples of S. guamare sp. n. were found with few specimens of S. nanella and S. wilsoni , and one of L. rayrobertsi . At other depths the concentrations were lower. S. nanella was common in the material examined, this species occupying a great extension of the bay. The other species were found concentrated in small areas of Guamaré Bay.
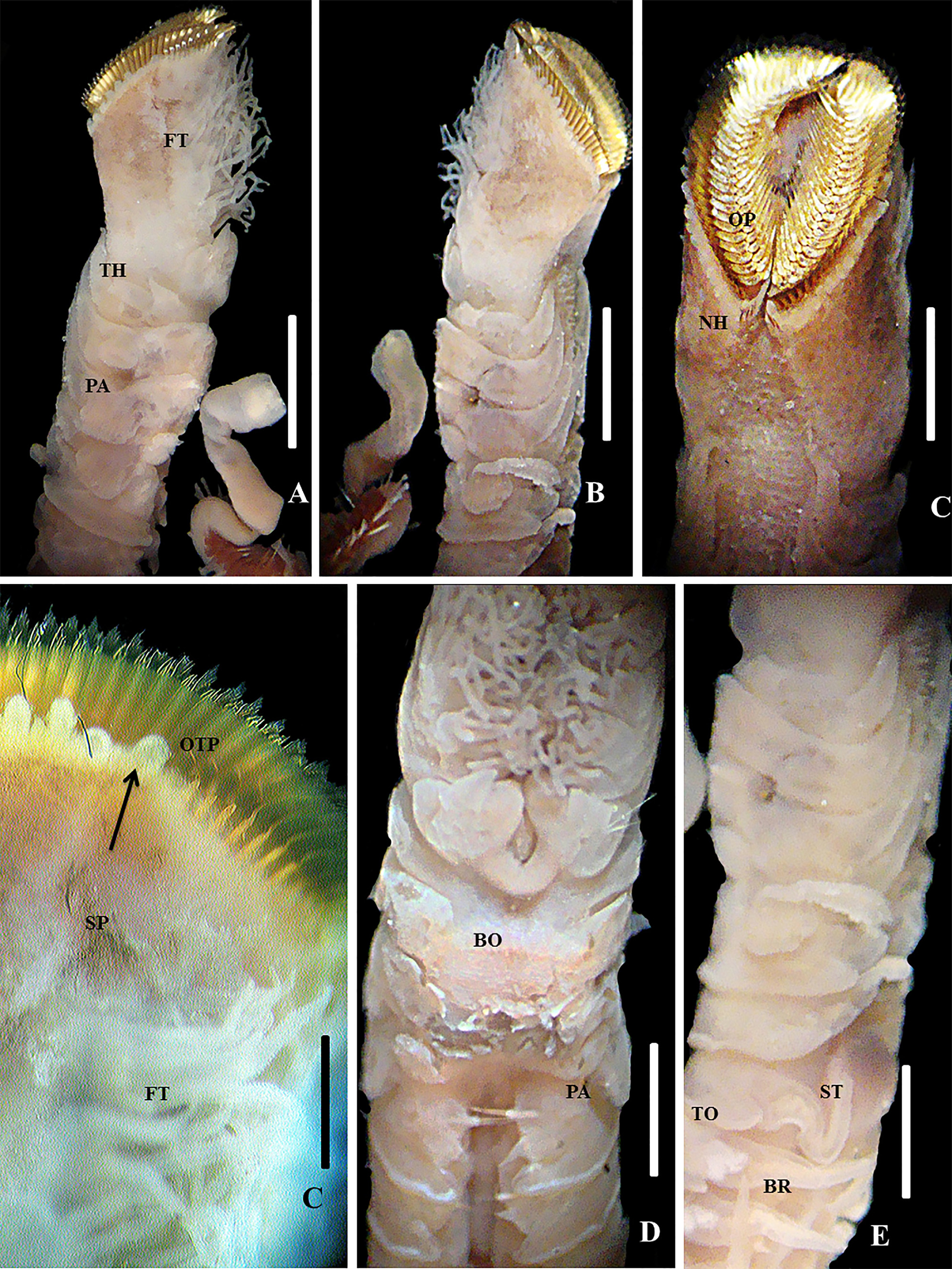
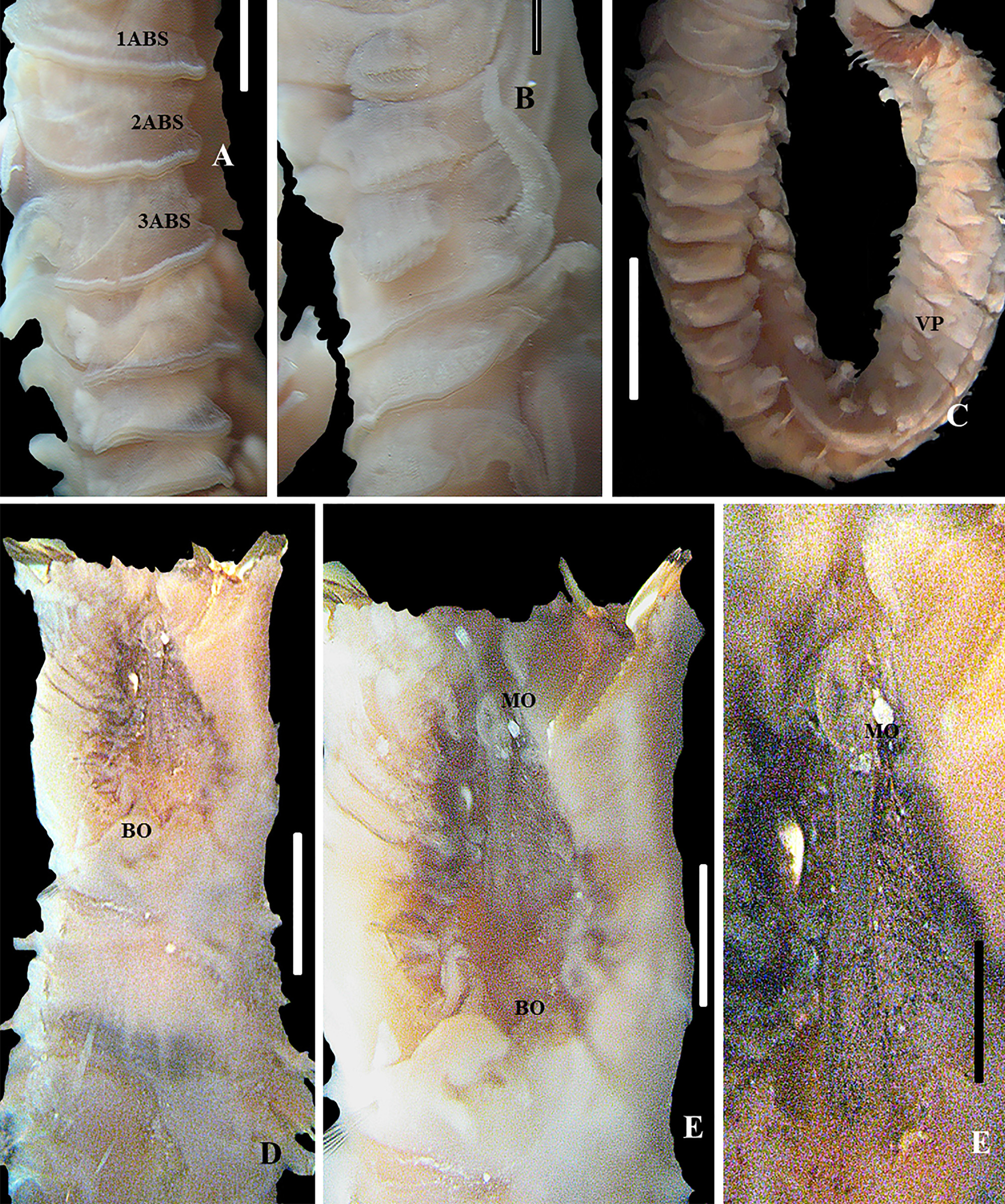


TABLE 1. Variation in morphological features of S. guamare n. sp. type series.
| Diameter of | Peduncle (length x width Parathoracic region | (length | x width | Abdomen width | Number of | Body length (mm) | Additional data | |
|---|---|---|---|---|---|---|---|---|
| operculum (mm) | (mm)) (mm)) | (mm) | abdominal segments | |||||
| Holotype | 3.0 | 5.0x3.0 2.5x2.0 | 2.0 | 40 | 55.0 | complete spec. | ||
| Paratype1 | 2.5 | 2.5x2.5 2.0x2.0 | 2.5 | 40 | 21.0 | complete spec. | ||
| Paratype 2 | 2.0 | 2.5x2.0 2.0x1.5 | 1.5 | - | - | incomplete spec. | ||
| Paratype 3 | 1.5 | 3.0x1.5 1.5x1.5 | 1.5 | - | - | incomplete spec. | ||
| Paratype 4 | 2.0 | 4.0x2.0 1.0x1.0 | 1.0 | 30 | 18.0 | incomplete spec. | ||
| Paratype 5 | 2.0 | 4.0x2.0 2.0x1.5 | 1.5 | 30 | 17.0 | incomplete spec. | ||
| Paratype 6 | 2.5 | 3.0x2.5 1.5x2.0 | 2.0 | - | incomplete spec. | |||
| Paratype 7 | - | 2.0x2.0 1.5x1.5 | 1.5 | 24 | 19.0 | incomplete spec. | ||
| Paratype 8 | 1.5 | 3.0x1.5 2.0x1.5 | 1.5 | 40 | 8.00 | complete spec. | ||
| Paratype 9 | 2.5 | 2.5x2.5 1.5x2.0 | 2.0 | 40 | 20.0 | complete spec. | ||
| Paratype 10 | 2.0 | 2.0x2.0 1.0x2.0 | 2.0 | 4 | 10.0 | incomplete spec. | ||
| Paratype 11 | 2.5 | 3.0x2.5 2.0x2.0 | 2.0 | 17 | 13.0 | incomplete spec. | ||
| Paratype 12 | - | 3.0x2.0 1.5x2.0 | 2.0 | - | incomplete spec. | |||
| Paratype 13 | - | 2.5x1.5 - | - | - | incomplete spec. | |||
| Paratype 14 | 1.0 | 1.5x1.0 1.5x1.0 | 1.0 | 23 | 20.0 | incomplete spec. | ||
| Paratype 15 | 3.0 | 5.0x3.0 2.5x2.5 | 2.5 | 4 | 6.0 | incomplete spec. | ||
| Paratype 16 | 2.0 | 3.0x2.0 2.0x1.5 | 1.5 | 5 | 7.0 | incomplete spec. | ||
| Paratype 17 | 1.0 | 2.0x1.0 1.0x1.0 | 1.0 | 24 | 19.0 | incomplete spec. | ||
| Paratype 18 | 1.0 | 2.0x1.0 1.0x1.0 | 1.0 | 24 | 19.0 | incomplete spec. | ||
| Paratype 19 | 1.0 | 2.0x1.0 1.0x1.0 | 1.0 | 7 | 8.0 | incomplete spec. | ||
| Paratype 20 | 1.0 | 2.0x1.0 1.0x1.0 | 1.0 | 9 | 8.0 | incomplete spec. | ||
| Paratype 21 | 2.0 | 3.0x2.0 2.5x2.5 | 2.0 | 1 | 8.0 | incomplete spec. |
TABLE 2. Alveolate species of Sabellaria that have three kinds of paleae.
| S. alveolata (Linnaeus, 1767) | present | 4–6 distal teeth lateral to a plume | deep excavation, concave, reflected outward | alveolate, geniculate, and concave | British Isles | Europe, Mediterranean, Western Africa | Kirtley, 1994; This study |
|---|---|---|---|---|---|---|---|
| S. bellis Hansen, 1882 View in CoL | present | 2–3 teeth in each side of plume | alveolate, light excavation, concave | geniculate, blade thicker, tip rounded | Rio de Janeiro, Brazil | Brazil to Argentina | Kirtley, 1994; This study |
| S. fissendis Grube, 1870 | not observed | 3–4 distal teeth lateral to a plume | alveolate, light excavation, concave | alveolate, geniculate, vertically pointed | Chile | Chile | Kirtley, 1994 |
| S guamarei sp. n. | present | 2–3 teeth in each side of plume | alveolate, light excavation, concave | geniculate blade thicker, tip rounded | Guamaré Bay | Guamaré Bay | This study |
| S. grueti Kirtley, 1994 View in CoL | present | 3–4 distal teeth lateral to a plume | alveolate, concave | geniculate, elongate | Burma, Andaman Sea | Andaman Sea | Kirtley, 1994 |
| S. ishikawi (Okuda, 1938) | present | 6–7 distal teeth | alveolate, concave | geniculate, elongate | San Thomé, Gulf of Guinea | West Africa | Kirtley, 1994 |
| S. javanica Auguener, 1934 View in CoL | present | a single apical sigmoid tooth | alveolate, light excavation, concave | geniculate, elongate | Soerabaia | Java | Kirtley, 1994 |
| S. moorei (Monro, 1933) View in CoL | present | 2–3 teeth in each side of plume | alveolate, light excavation, concave | geniculate blade thicker, tip rounded | Panama | Panama | Kirtley, 1994; This study |
| S. myriansis Parab & Gaikward, 1990 | not observed | 3–4 teeth in each side of plume, tip curved | alveolate, light excavation, concave | distal tip geniculate, elongate blade | West coast of India | West coast of India | Kirtley, 1994 |
| DISCUSSION |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Sabellaria guamare
| Santos, André Souza Dos, Brasil, Ana Claudia Dos Santos & Christoffersen, Martin Lindsey 2014 |
S. grueti
| Kirtley 1994 |
S. myriansis
| Parab & Gaikward 1990 |
S. wilsoni
| Lana and Gruet 1989 |
S. ishikawi
| Okuda 1938 |
S. javanica
| Auguener 1934 |
S. moorei
| Monro 1933 |
S. nanela
| Chamberlin 1919 |
S. bellis
| Hansen 1882 |
S. fissendis
| Grube 1870 |
S. alveolata
| Linnaeus 1767 |