
Namanereis, Glasby & Fiege & Damme, 2014
|
publication ID |
https://doi.org/10.1111/zoj.12130 |
|
DOI |
https://doi.org/10.5281/zenodo.10541948 |
|
persistent identifier |
https://treatment.plazi.org/id/039D87CE-197B-FFA6-2663-F9D8D19873BD |
|
treatment provided by |
Marcus |
|
scientific name |
Namanereis |
| status |
|
HABITATS BY NAMANEREIS View in CoL
In general, the colonization of subterranean habitats by stygobitic taxa from marine or freshwater epigean ancestors can occur in different ways and often at different times. These often ancient (Gondwanan) colonizations and fragmentations in stygobitic taxa lead to a number of current biogeographical disjunctions (e.g. Tethyan distribution patterns) explained by vicariances (e.g. crustaceans; Stock, 1993). In crustacean isopods, Wägele (1990) reviewed two colonization pathways for stygobitic isopods, one from coastal groundwater and one from surface waters. Holsinger (1994) sug- gested three pathways for amphipods, one from epigean freshwater ancestors (limnostygobionts) and two possible scenarios for marine/brackish ancestors (thalassostygobionts) by marine regressions and land uplift or adaptive shifts during fluctuating sea levels. The latter scenario is linked with a Tethyan colonization ( Holsinger, 1994). In stygobitic copepods, also a group of marine origin, several lineages colonized groundwater independently as a result of multiple invasions; several went through an epigean freshwater phase before entering groundwater (e.g. Galassi, 2001), but several colonized directly from the sea ( Boxshall & Jaume, 2000; Galassi et al., 2009). What is the scenario for this particular lineage of polychaetes?
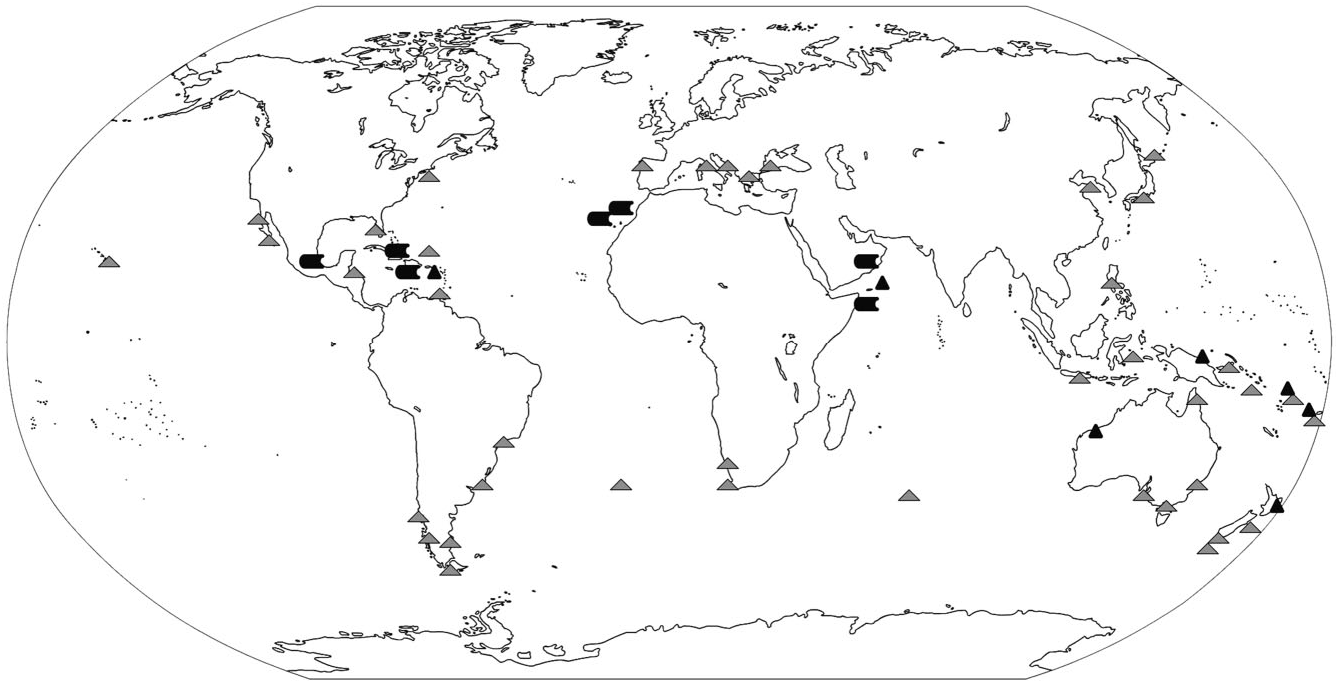
Although some of the similarities amongst groundwater species may be attributable to convergence related to adaptation to a similar habitat, the major morphological difference (viz. bifid vs serrated jaws; Table 1) between groundwater species might have an historical explanation. The Arabian region including Socotra (derived from the Afro-Arabian Plate) and the Caribbean have the highest diversity of groundwater namanereids in the world. The bifid-jawed species are found at disjunct localities, from Socotra Island westward to the Caribbean and Central America; by comparison, the serrated-jawed species are more widely distributed from the Caribbean ( Sint Eustatius and Jamaica) through Socotra (Abd al Kuri) eastward to Fiji and New Zealand ( Fig. 8 View Figure 8 ; Table 1). It seems therefore that the two differently jawed species living on the Socotra Archipelago may not be closely related: the serrated-jawed forms, with the exception of N. sublittoralis , were all connected by a relatively short length of Tethyan coastline in the Late Jurassic just prior to the breakaway of Gondwana from Pangaea, so it is conceivable that a single colonization event occurred, followed by speciation as a result of the fragmentation of Gondwana. By comparison, the Afro- Caribbean distributed bifid-jawed species appear to be more recent, possibly Cretaceous. At this time the Proto- Caribbean Sea was still connected with the Tethys Sea, allowing for the possibility of a widespread marine ancestor – from north-east Africa to the western Caribbean – to invade the land in a second colonization event. Speciation may have occurred within the bifid-jawed group around the time of the closure of Tethys Sea with the collision of African and Arabian Plates with Eurasia during the Miocene about 20 Mya [e.g. Glasby, (2005)]. This also corresponded with the emergence of the Canary Islands. Cenozoic speciation of the bifid-jawed species would correspond to geological links in Socotra – separation between N. socotrensis sp. nov. from the northern coast of Socotra Island and N. araps from Oman, which seem to share a common ancestor, corresponds with the original limestone deposits in which these animals are found. Until the Miocene ( c. 18 Mya), Socotra was not an island but geologically part of southern Oman ( Dhofar Region), and both regions have the same large Late Cretaceous/ Palaeocene/Eocene karstic limestone deposits until then and most areas were under shallow epicontinental seas until the end of the Eocene (see Cheung & DeVantier, 2006 for overview and references). In fact, it is the same suite of tectonic events that caused the closure of the Tethys Sea, which separated the Socotra Archipelago from the mainland and resulted in uplift of the northern and southern flanks of the Gulf of Aden.
The Gondwanan group includes the new species from Pilbara, north-west Australia. The Pilbara is thought to be very old, having remained more or less emergent since the Proterozoic (> 545 Mya); it is home for several ancient (probably endemic) freshwater lineages of amphipods, isopods, copepods, and ostracods, which do not occur in the adjacent regions that were inundated by the sea in the Cretaceous and Eocene ( Eberhard, Halse & Humphreys, 2005; Humphries, 2008). The new species of Namanereis from the Pilbara is likely to belong to this same ancient assemblage. It is important to note that the step from marine to freshwater in Namanereis was taken through the subterraneous route by more than half of the species (12 out of 17), although not all. The diversity in ecology in non-groundwater species ( Glasby, 1999; Williams, 2004), ranging from the intertidal zone to leaf-litter, suggests that the colonization of land might not have been one simple event with subsequent radiation. We should note here that for such ancient (e.g. Mesozoic) groups, the distribution in terrestrial aquatic ecosystems may have been very different from today, as continental aquatic habitats were completely different (e.g. much more oligotrophic) and only became similar to those we know today near the end of the Palaeogene in the Cenozoic (e.g. Ponomarenko, 1996). It means that some species might have had wider distributions in epigean oligotrophic systems, and that the hyporheic or karstic fissures could also function as refuge habitats.
Perhaps a colonization history of Namanereis can be distilled from the range of current habitats: from the littoral/intertidal zone, through entrapment into land-locked fluctuating brackish-water/freshwater systems (e.g. freshwater−brackish-water interface in coastal caves in karstic regions), in analogy with thalassostygobitic amphipods and copepods (cf. Holsinger, 1994; Galassi, 2001). Such a scenario could explain the presence of Namanereis in caves at higher elevations, such as N. gesae in a cave at 700 m on Abd al Kuri, or N. cavernicola in Mexico, which were originally at sea level, yet uplifted later. This scenario is plausible for Namanereis species occupying orogenic (uplifted) terranes. The latter would imply a relatively ‘direct’ initial colonization, and then further colonization of other habitats by run-off. An alternative hypothesis, or possibly a second parallel scenario, would be, as in several stygobitic crustaceans, that these polychaetes entered the groundwater in two steps: from marine environments to epigean freshwater habitats from fluctuating brackish/freshwater coastal environments such as coastal lagoons or mangroves and later taking refuge in groundwater (hyporheic, caves) through run-off. The scenario is plausible for species occupying ancient cratons, like the two new species described here. Again, such scenarios are speculative and perhaps hard to build on biogeography, ecology, and morphology, yet the current lack of a molecular framework in Namanereis should not be an obstacle for exploring hypotheses on their colonization history and evolution. Whether the venture into freshwater and anchialine habitats by Namanereis happened in a single colonization event that pre-dates the break-up of Gondwana ( Glasby & Timm, 2008) or involved at least two independent events separated by millions of years (present study) remains unanswered. The two hypotheses present very different scenarios for evolution within the group, and could presumably be tested by application of molecular clock methods.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.