
Orobdella octonaria Oka, 1895
|
publication ID |
https://doi.org/ 10.12782/sd.17.2.227 |
|
publication LSID |
lsid:zoobank.org:pub:3AEBB603-BDC5-4518-85BF-92FFFDE10739 |
|
persistent identifier |
https://treatment.plazi.org/id/03A287F4-FFFC-FFAE-FED8-2AD1DAF6FA53 |
|
treatment provided by |
Felipe |
|
scientific name |
Orobdella octonaria Oka, 1895 |
| status |
|
urn:lsid:zoobank.org:act:BFBDD5FB-FCC1-470F-A93C-1397F5576C9B ( Figs 2–7 View Fig View Fig View Fig View Fig View Fig )
Orobdella octonaria Oka, 1895: 286–288 , pl. 28, figs 3, 11– 14, pl. 29, fig. C; Oka 1910a: 61; Oka 1910b: 178; Oka and Nagao 1965: 574; Soós 1966: 381, 397; Lukin 1976: 466– 467; Sawyer 1986: 680, 747.
Kumabdella octonaria: Richardson 1971: 591–599 , figs 1–3.
Diagnosis. Somite VII ventrally septannulate (octannulate dorsally). Somites VIII–XXV complete octannulate. Gastropore conspicuous at XIII b2. Gastroporal duct slightly winding at junction with gastropore, tubular but slightly bulbous at junction with crop. Male gonopore at XI c11. Female gonopore inconspicuous at XIII b2. Gonopores separated by 1/2 or 1/4+10+1/2 or 3/4 annuli.
Lectotype designation. One syntype, NSMT-An 415, deposited in the NSMT was chosen as the lectotype for O. octonaria in this study, since the locality of the specimen is indicated in detail and the syntype is dissectable. NSMT-An 415 was found dissected dorsally from somite XVII c9 to the caudal sucker ( Fig. 2A View Fig ), and labeled as ‘type’. The specimen was collected from Mt. Yusakayama, Hakone, Kanagawa Prefecture, Japan.
Material examined. Lectotype: NSMT-An 415, a mature specimen of 221.8 mm in BL, dissected, collected from Mt. Yusakayama (coordinates of the crest: 35°13′41″N, 139°04′46″E), Hakone, Kanagawa Prefecture, Japan, on 1 June 1894 GoogleMaps . Additional material: two specimens collected from under rocks along a mountain trail at Mt. Sengenyama, Hakone , Kanagawa Prefecture, Japan, by Takafumi Nakano on 7 July 2011: KUZ Z181 View Materials (alt. 585 m, 35°14′03″N, 139°04′12″E), dissected, and Z182 (alt. 577 m, 35°14′04″N, 139°04′13″E) GoogleMaps .
Description of lectotype. Body firm, muscular, elongated, gaining regularly in width in caudal direction, dorsoventrally depressed, sides nearly parallel from midlength to point just anterior of caudal sucker, BL 221.8 mm, BW 15.0 mm ( Fig. 2A, B View Fig ). Caudal sucker ventral, oval, its diameter slightly less than BW ( Fig. 2B View Fig ). Color in life unknown. Color faded in preservative, without any dark lines.
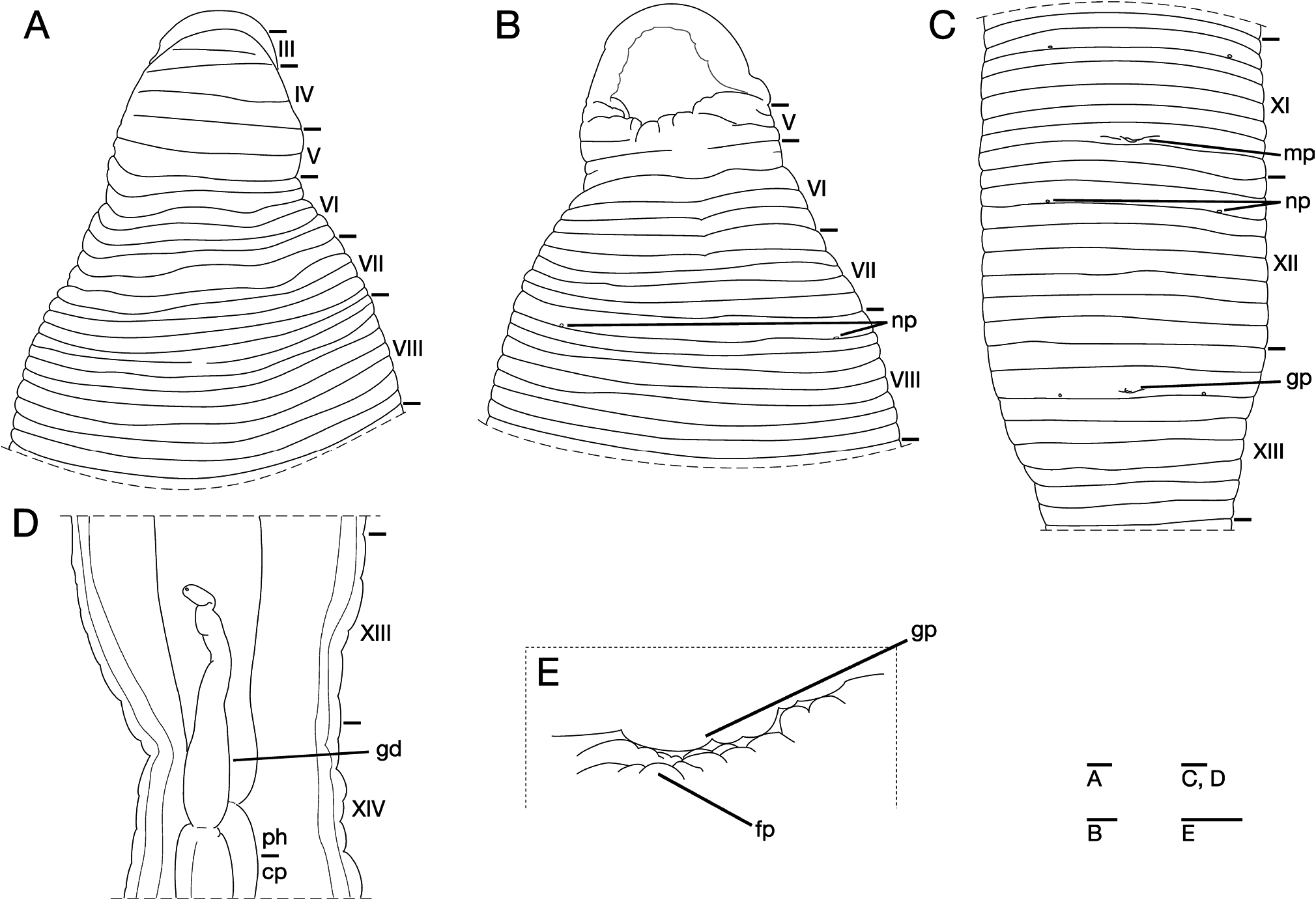
Somite I completely merged with prostomium ( Fig. 3A View Fig ). Somite II uniannulate. Somites III–V biannulate ( Fig. 3A, B View Fig ), V a3 forming posterior margin of oral sucker ( Fig. 3B View Fig ). Somite VI ventrally quadrannulate (sexannulate dorsally), a1 (b1, b2 dorsally) = a2 (b3, b4 dorsally)>b5=b6; a1 and a2 with obvious furrow dorsally ( Fig. 3A, B View Fig ). Somite VII quinquannulate, a1>b3=b4=b5=b6 ( Fig. 3A, B View Fig ). Somite VIII ventrally septannulate (octannulate dorsally), a1 (b1, b2 dorsally)>b3=b4=c9=c10=c11=c12; a1 with obvious furrow dorsally ( Fig. 3A, B View Fig ). Somites IX–XXVI octannulate, b1=b2=b3=b4=c9=c10=c11=c12 ( Fig. 3C View Fig ). Annulation of somite XXVII undecidable, comprised of one annulus (169th) with several slight furrows, 169th annulus being last complete annulus on venter; anus behind it with no postanal annulus.
Anterior ganglionic mass in VI a1. Ganglion VII in a1. Ganglia VIII–XI in b3 and b4 of each somite. Ganglion XII in b3. Ganglion XIII in b4 and c9. Ganglia XIV–XIX in b3 of each somite. Ganglia XX–XXV undetectable. Ganglion XXVI in b4. Posterior ganglionic mass in XXVII.
Eyes not detectable. Nephridiopores in 17 pairs, ventrally at posterior margin of VIII a1, and at posterior margin of b2 of each somite of IX–XXIV ( Fig. 3B, C View Fig ). Papillae numerous, minute, hardly visible, one row on every annulus.
Pharynx agnathous, euthylaematous, reaching to XIV c9/ c10. Crop tubular, acecate, sphincter between crop and intestine undetectable. Gastropore conspicuous, ventral, at posterior of XIII b2 ( Fig. 3C, E View Fig ). Gastroporal duct muscular, slightly winding at junction with gastropore, tubular but slightly bulbous at junction with crop, joining with crop in XIV b4 ( Fig. 3D View Fig ). Intestine and rectum undetectable.
Male gonopore at posterior of XI c11 ( Fig. 3C View Fig ). Female gonopore at posterior of XIII b2, inconspicuous, located diagonally behind gastropore ( Fig. 3E View Fig ). Gonopores separated by 1/4+10+3/4 annuli ( Fig. 3C View Fig ). Testisacs undetectable. Paired epididymides in XVII b1/b2 to XIX b2 ( Fig. 4A View Fig ). Ejaculatory bulbs absent. Ejaculatory ducts in XI c9 to XVII b1/b2, loosely coiled, each widening from junction with epididymis, narrowing at junction with atrial cornu, then turning gradually inward toward atrial cornu without pre-atrial loop ( Fig. 4A–D View Fig ). Pair of atrial cornua in XI c9 to c11, muscular, fusiform ( Fig. 4B–D View Fig ). Atrium body short, muscular, globular in XI C10 and c11 ( Fig. 4A–D View Fig ). Penis sheath and penis absent. Ovisacs one pair, thin-walled, globular, in XIII b3 and b4 ( Fig. 4A, E View Fig ). Oviducts thin-walled, right oviduct crossing ventrally beneath nerve cord, both oviducts converging into common oviduct in XIII b3 ( Fig. 4A, E View Fig ). Common oviduct thin-walled, short, directly descending to female gonopore ( Fig. 4E View Fig ).
Description of other specimens. Maximum BL 132.7 mm, maximum BW 7.71 mm ( Fig. 5A, B View Fig ). In life, dorsal surface bluish gray without any dark lines ( Fig. 6 View Fig ), ventral surface bluish white. Color faded in preservative.
Somite VI ventrally quadrannulate (sexannulate dorsally) (KUZ Z181) ( Fig. 7A, B), or ventrally triannulate (quinquannulate dorsally), a1 (b1, b2 dorsally)=a2 (b3, b4 dorsally)>a3 (KUZ Z182). Somite VII ventrally quiquannulate (sexannulate dorsally), a1 (b1, b2 dorsally)>b3=b4= b5=b6; a1 with obvious furrow dorsally ( Fig. 7A, B). Somite XXVI octannulate, b1=b2=b3=b4=c9=c10=c11= c12, c12 being last complete annulus on venter (KUZ Z181) ( Fig. 7C, D), or sexannulate, annulation formula undecidable, composed of six (160th–165th) annuli, 165th annulus being last complete annulus on venter (KUZ Z182). Somite XXVII biannulate (KUZ Z182), or probably triannulate, a1>a2>a3; a1 with slight furrow dorsally (KUZ Z181) ( Fig. 7C, D).
Anterior ganglionic mass in VI a2 and b5. Ganglia VIII–X, XII, XIV–XXI in b3 and b4 of each somite. Ganglia XI and XIII in b4 of each somite. Ganglia XXI–XXV in b3 of each somite. Ganglion XXVI in b1 and b2. Posterior ganglionic mass in XXVI c9–c11.
Eyes three pairs, first pair dorsally on posterior margin of II, second and third pairs dorsolaterally on posterior margin of V (a1+a2) ( Fig. 7A).
Pharynx reaching to XIV c9. Crop in XIV c9 to XXI c12. Gastropore at middle of XIII b2 ( Fig. 7E). Gastroporal duct joining with crop in XIV b2/b3. Intestine tubular, acecate, in XXI c12 to XXV b1. Rectum tubular, thin-walled.
Male gonopore at middle of XI c11. Female gonopore at middle of XIII b2. Gonopores separated by 1/2+10+1/2 annuli. Paired epididymides in XVII b4/c9 to XIX c9/c10. Ejaculatory ducts in XI c9 to XVII b4/c9.
Distribution. Known from mountainous regions of Honshu, Shikoku, and Kyushu, Japan ( Oka and Nagao 1965).
Remarks. Orobdella octonaria is easily distinguished from the other eight species of Orobdella in having midbody somites that are octannulate. Five species, O. esulcata Nakano, 2010 , O. kawakatsuorum Richardson, 1975 , O. koikei Nakano, 2012 , O. tsushimensis Nakano, 2011 , and O. whitmani , have quadrannulate mid-body somites. Three species, O. dolichopharynx Nakano, 2011 , O. ijimai , and O. shimadae Nakano, 2011 , have mid-body somites that are sexannulate. Octannulate species other than O. octonaria have not been reported yet.
Richardson (1971) redescribed of O. octonaria based on a single specimen collected from Nagano Prefecture, Japan (NSMT-An 52). His specimen could not be reexamined in this study, since it was not found in the annelid collection of the NSMT. However, several points of Richardson’s (1971) description are discordant with the present study. First, he noted that the female gonopore is at XIII b2 but not positioned close to the gastropore (XIII c1/c 2 in his paper, i.e., XII/XIII in current terminology). In the lectotype and the newly obtained specimens, in contrast, the female gonopore is close to the gastropore and both pores open in the middle of XIII b2. In his description of O. kawakatsuorum, Richardson (1975b) mentioned that the female gonopore and gastropore of that species as well are not together. In actuality, these two pores are closely adjacent in the holotype and the newly obtained specimens (Nakano 2012). In addition, the female gonopore in all the other gastroporous species of Orobdella is also close to the gastropore ( Nakano 2010, 2011b, c, 2012). These facts suggest that Richardson (1971, 1975b) misjudged the positions of the female gonopores and gastropores in the species of Orobdella he studied. Richardson (1971) also noted that O. octonaria possesses a pseudopore in XIII b4/b5 (i.e., b4/c 9 in current terminology), but such a pore was not detected in the specimens examined in the present study. As for the genital organs, Richardson (1971) noted that a male atrium does not exist, and that the two oviducts do not converge into a common oviduct but descend directly and separately to the female gonopore, however, O. octonaria actually does have a developed male atrium ( Fig. 4B–D View Fig ) and a common oviduct ( Fig. 4E View Fig ).
Richardson (1971) established the genus Kumabdella Richardson, 1971 for O. octonaria , diagnosed principally by its octannulate mid-body somites. He also suggested that another new genus should be erected for sexannulate O. ijimai . He attributed great weight to the mid-body somite annulation in leech classification, and split many genera on this basis (e.g., Richardson 1969, 1975a). Recent molecular phylogenetic studies have shown that mid-body somite annulation does not reflect phylogenetic relationships in erpobdellid ( Trontelj and Sket 2000) and haemadipsid leeches ( Borda and Siddall 2011). This suggests that the generic classification on the basis of mid-body somite annulation should be reconsidered in other families as well. In the case of Orobdella , Richadson’s (1971) classification has not been followed by subsequent taxonomic workers ( Lukin 1976; Sawyer 1986; Nakano 2011c; Oceguera-Figueroa et al. 2011; Nakano et al. 2012). Molecular phylogenetic analyses have revealed that O. octonaria is included in a clade of mostly quadrannulate species, and that sexannulation and octannulation evolved in parallel in Orobdella (Nakano 2012; Nakano et al. 2012). Inasmuch as O. octonaria is clearly in accord with the diagnostic characters of Orobdella as modified by Nakano (2011c), Kumabdella should indeed be regarded as a junior synonym of Orobdella , in accordance with the five post-1971 works cited above.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Orobdella octonaria Oka, 1895
| Nakano, Takafumi 2012 |
Kumabdella octonaria: Richardson 1971: 591–599
| Richardson, L. R. 1971: 599 |
Orobdella octonaria
| Sawyer, R. T. 1986: 680 |
| Lukin, E. I. 1976: 466 |
| Soos, A. 1966: 381 |
| Oka, A. & Nagao, Z. 1965: 574 |
| Oka, A. 1910: 61 |
| Oka, A. 1910: 178 |
| Oka, A. 1895: 288 |