
Myriowenia undetermined
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2012.00850.x |
|
persistent identifier |
https://treatment.plazi.org/id/03AE8729-AB04-D44A-FED4-FCBDA7DDF89F |
|
treatment provided by |
Marcus |
|
scientific name |
Myriowenia undetermined |
| status |
|
FIGURES 9F–H View Figure 9 , 12 View Figure 12 AND 13 View Figure 13 , TABLE 6
Material examined (Appendix 1): New South Wales (128 specs), south of Sydney: Cobblers, Wattamolla , Bass Point , Cape Banks and Marley , in sandy bottoms, 40–70 m.
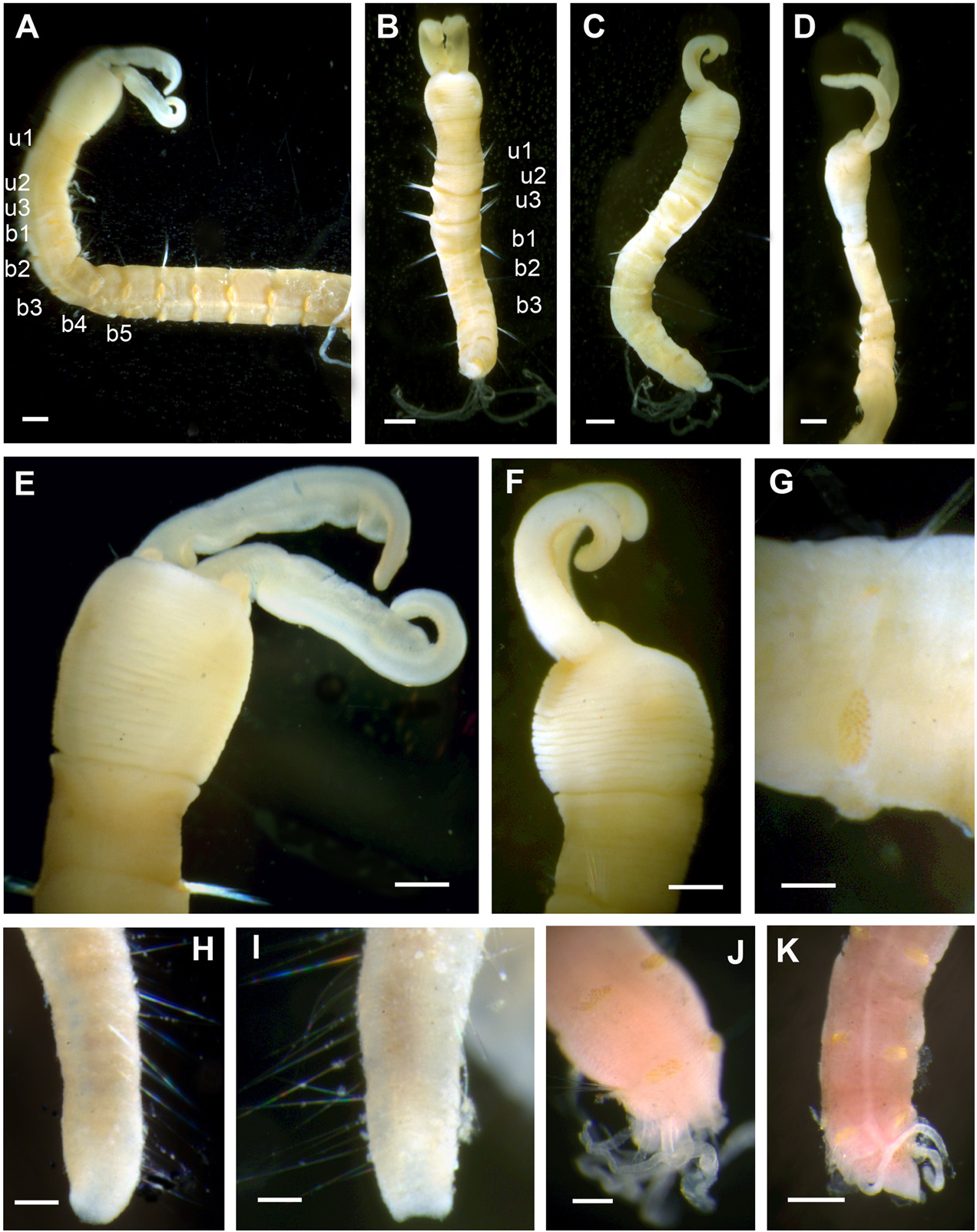
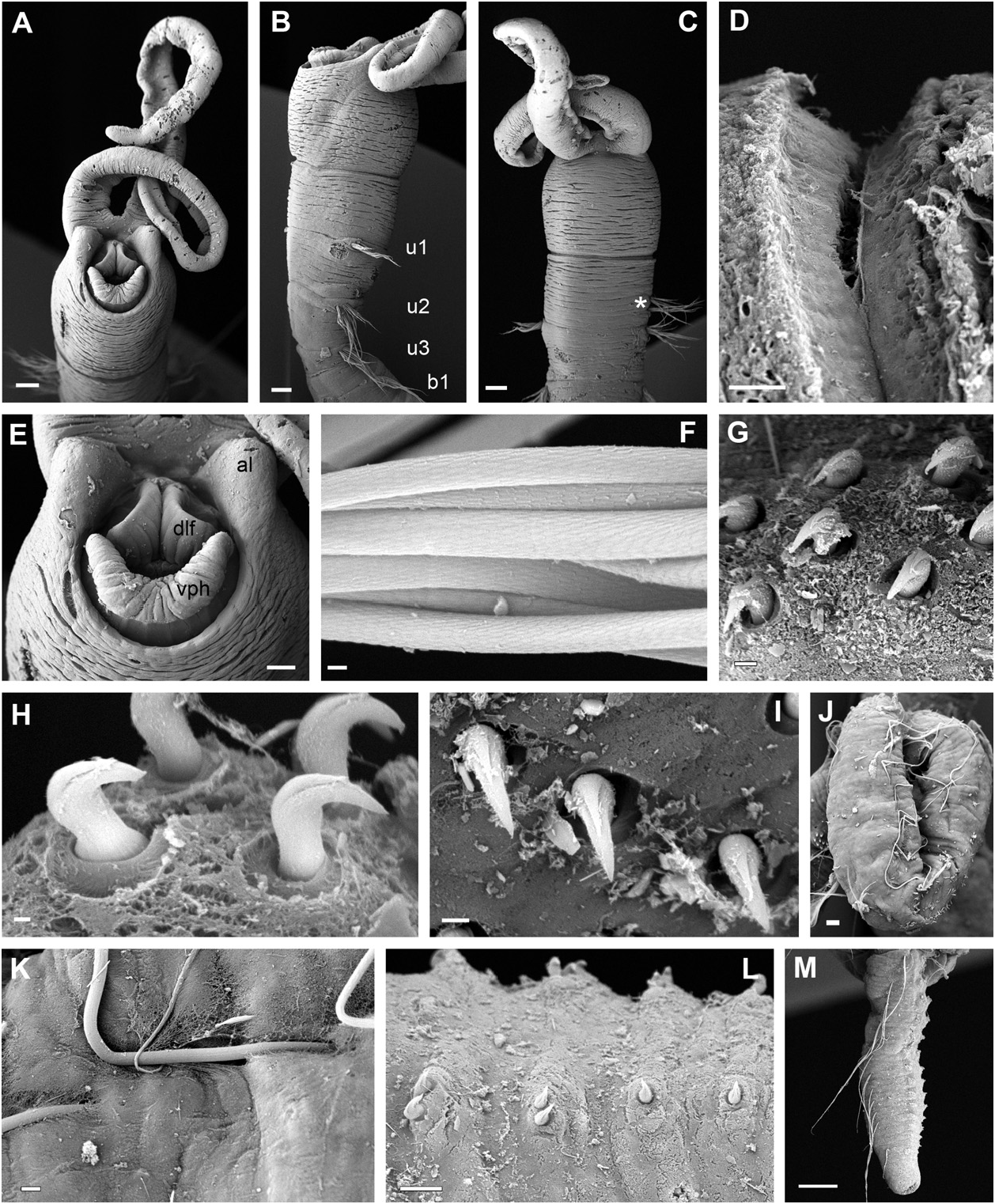
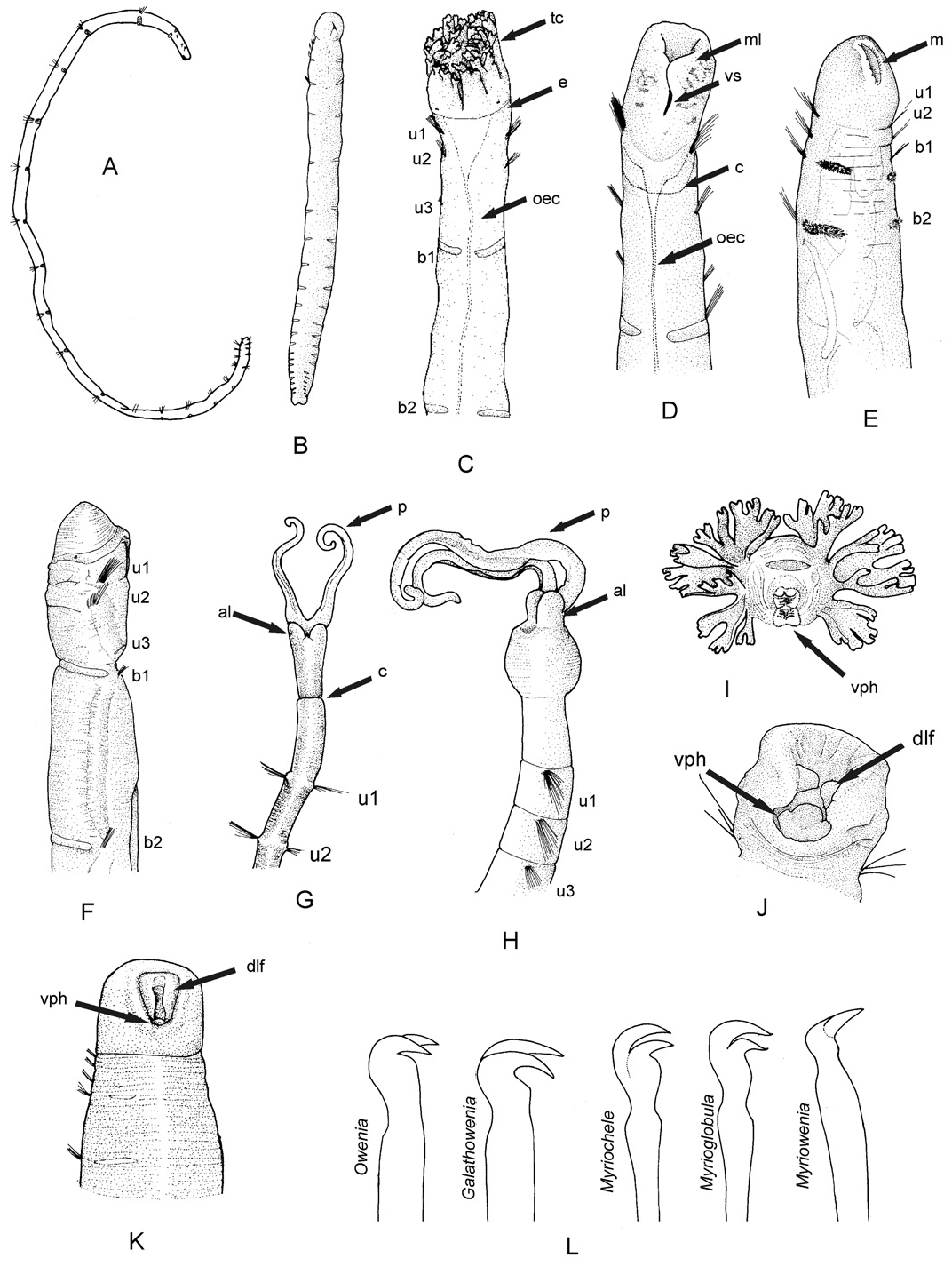
Description: Largest complete specimen measuring 14 mm long and 0.8 mm wide, with 55 chaetigers. Body cylindrical in cross section ( Fig. 12A–C View Figure 12 ), with slightly tapering posterior end. Epithelium wrinkled in head and anterior segments. No longitudinal ciliated grooves present. Nervous system evident as white ventral longitudinal band. Head region rounded, wider than rest of the body in most specimens, distinctly set off from segmented region by constriction ( Figs 9F, G View Figure 9 , 12A–F View Figure 12 , 13A–D View Figure 13 ). Two anterior grooved palps similar in length to uniramous region ( Figs 12A–F View Figure 12 , 13A–C View Figure 13 ), with rounded anterior lobes on each base (( Figs 12E View Figure 12 , 13A, E View Figure 13 ). Anteroventral mouth with ventral pharyngeal organ and dorsolateral fold present and exposed in some specimens ( Fig. 13A, E View Figure 13 ). Eyespots not observed. First three chaetigers uniramous ( Figs 12A, B View Figure 12 , 13B View Figure 13 ), similar in length or first one slightly longer ( Figs 12A, B View Figure 12 , 13B View Figure 13 ); biramous segments also similar in length ( Fig. 12A–C View Figure 12 ) to posterior end, becoming progressively shorter and thinner ( Figs 12H, I View Figure 12 , 13M View Figure 13 ). Notopodia with capillaries ( Fig. 12G View Figure 12 ) with proximal end smooth and distal end with scale covering ( Fig. 13F View Figure 13 ); diminishing in length ventrally within each fascicle. Acicular chaetae absent. Neuropodial tori narrow, with uncini arranged in two to three irregular rows in anterior and middle segments ( Fig. 13G View Figure 13 ) to one short row consisting of one or two uncini in posterior segments ( Fig. 13L View Figure 13 ). Uncini with two curved teeth, arranged one above the other, distal tooth smaller than proximal one ( Fig. 13G–I View Figure 13 ). Posterior end cylindrical, distally tapered, with compressed segments; pygidium as a rim encircling anus ( Figs 12H, I View Figure 12 , 13M View Figure 13 ). Tube very thin and flexible only covered by a few fine sand grains, not heavily cemented. Several specimens with posterior ends exhibiting regeneration processes at the broken posterior end of the body (( Figs 9H View Figure 9 , 12J–K View Figure 12 ). Colour of specimens pale yellow after preservation.
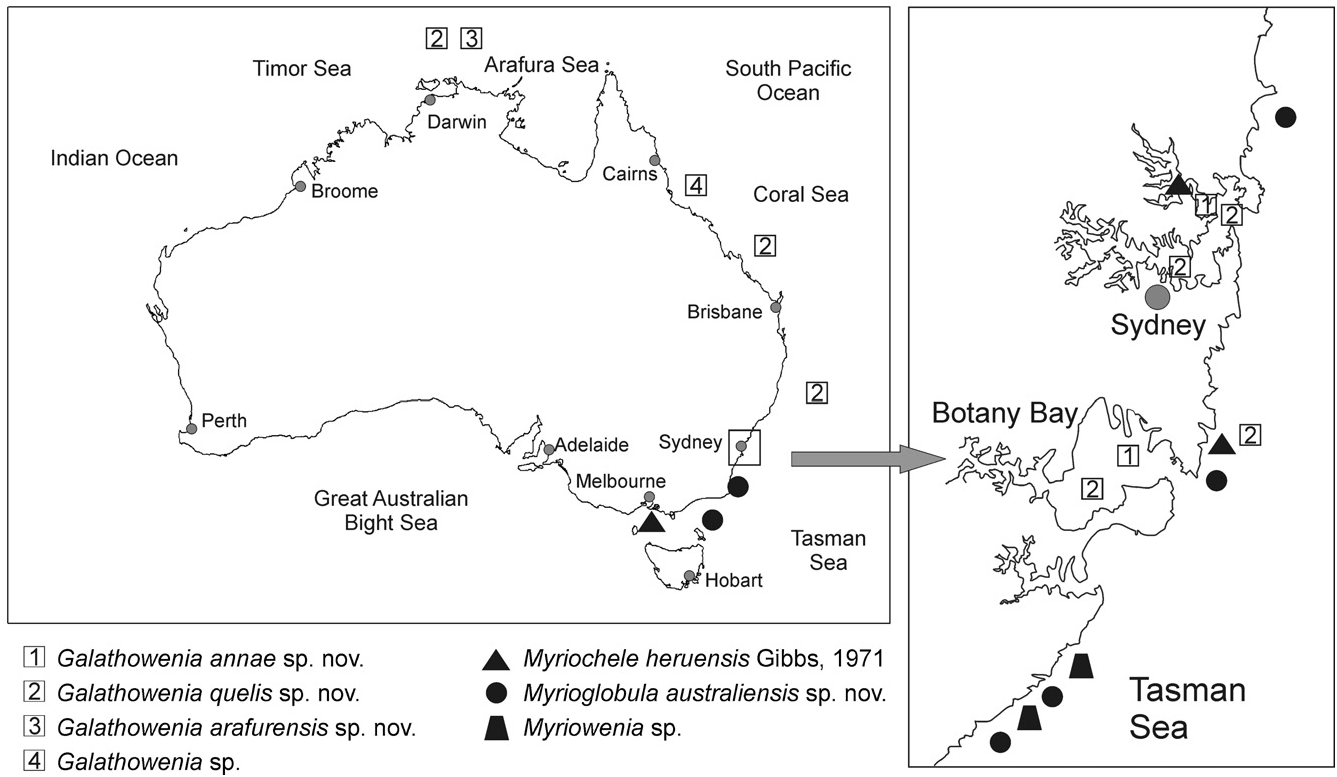
Ecological notes: Specimens found in sandy bottoms and at 40–70 m depths, in localities south of Sydney , New South Wales ( Fig. 14 View Figure 14 ) .
Remarks: Three species of Myriowenia have been described so far, Myriow. californiensis Hartman, 1960 , from California ( Fig. 15H View Figure 15 ), and two others from the Gulf of Mexico, Myriowenia gosnoldi Hartman, 1965 , ( Fig. 15G View Figure 15 ) and Myriowenia sp. A Milligan, 1984 (see Table 6). Differences amongst these species have not been addressed in previous studies because in most cases insufficient material was available or its condition was not optimal to check intraspecific variation. For example, what has been interpreted as a cylindrical head in Myriow. gosnoldi and Myriowenia sp. A ( Hartman, 1965; Milligan, 1984) has also been observed in some of the specimens from New South Wales (e.g. Fig. 12D View Figure 12 ) and could be because of the flexibility of this region, but certainly is not a diagnostic character of these species.
The presence of ‘a pair of pygidial cirri’ in Myriow. gosnoldi ( Hartman, 1965; 347: fig. 44b) resembles the regeneration processes that we have also observed in some broken fragments. Some Australian specimens of Myriowenia sp. , like other oweniids herein described, show a certain type of filamentous processes in the fragmented posterior body part. These processes resemble extensions of the body integument ( Fig. 12J, K View Figure 12 ), resulting in a different appearance of the posterior end when compared with complete (intact) specimens, with the posterior segments being compressed and slightly thinner towards the low rim pygidium ( Fig. 12H, I View Figure 12 ). Moreover, in some Australian specimens other processes appear to originate from the side pores of some midbody segments ( Fig. 13J, K View Figure 13 ). The true composition of these processes and their function are unknown and deserve more investigation. The main character that separates Myriowenia sp. and Myriow. gosnoldi or Myriowenia sp. A is the absence of a collar membrane between the cephalic and the segmented region (present in both Myriow. gosnoldi and Myriowenia sp. A ). Myriowenia sp. is therefore more similar to Myriow. californiensis (sensu Hartman, 1960 and Blake, 2000), in sharing *According to Blake (2000).
†The specimen described by Hartman (1965) is probably an anterior fragment undergoing regeneration.
‡Segment 2 according to author’s description, but not from drawings ( Milligan, 1984: fig. 46-4).
nd, no data available.
the presence of short uniramous segments. However, because the pygidium and posterior segments have not been described in Myriow. californiensis , we cannot compare these two, probably disjunct, Myriowenia species in detail.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |