
Myriochele australiensis, Capa & Parapar & Hutchings, 2012
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2012.00850.x |
|
persistent identifier |
https://treatment.plazi.org/id/03AE8729-AB0B-D44E-FCD9-F956A5B0F9EE |
|
treatment provided by |
Marcus |
|
scientific name |
Myriochele australiensis |
| status |
sp. nov. |
MYRIOCHELE AUSTRALIENSIS View in CoL SP. NOV.
( FIGURES 9E View Figure 9 , 10 View Figure 10 , 11 View Figure 11 , 14 View Figure 14 , TABLES 1, 5
Holotype: AM W20675, New South Wales, east of Long Reef , 33°44′43″S, 151°22′43″E, sand, 60 m, 13.iv.1989. GoogleMaps
Paratypes: AM W37827, same sample (42 specs) .
Other material examined (Appendix 1): New South Wales (175 specs): Malabar, Cobblers, Cape Banks, Wattamolla, Basspoint, Murramarang National Park ; Victoria (1 spec.), Bass Strait .
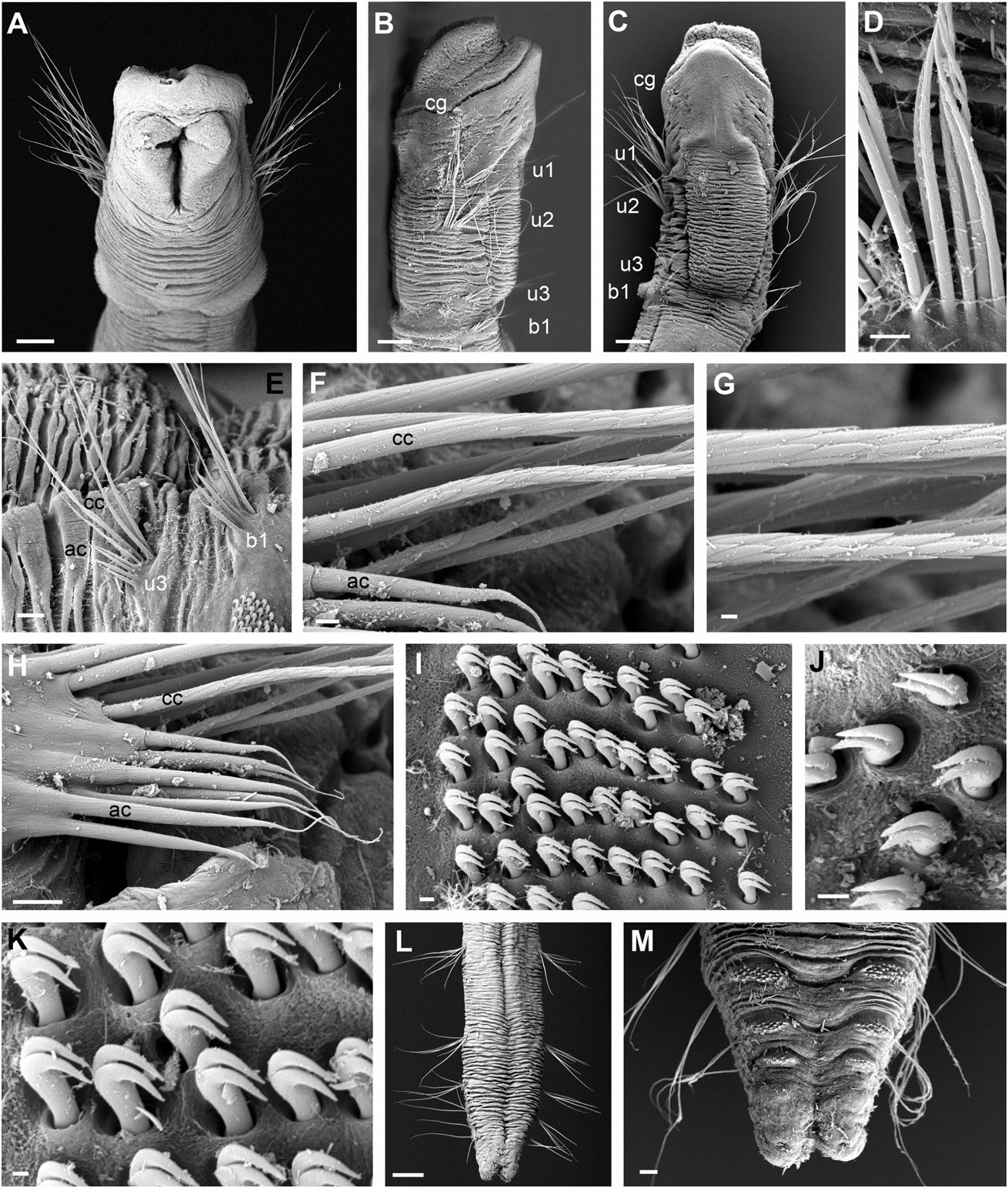
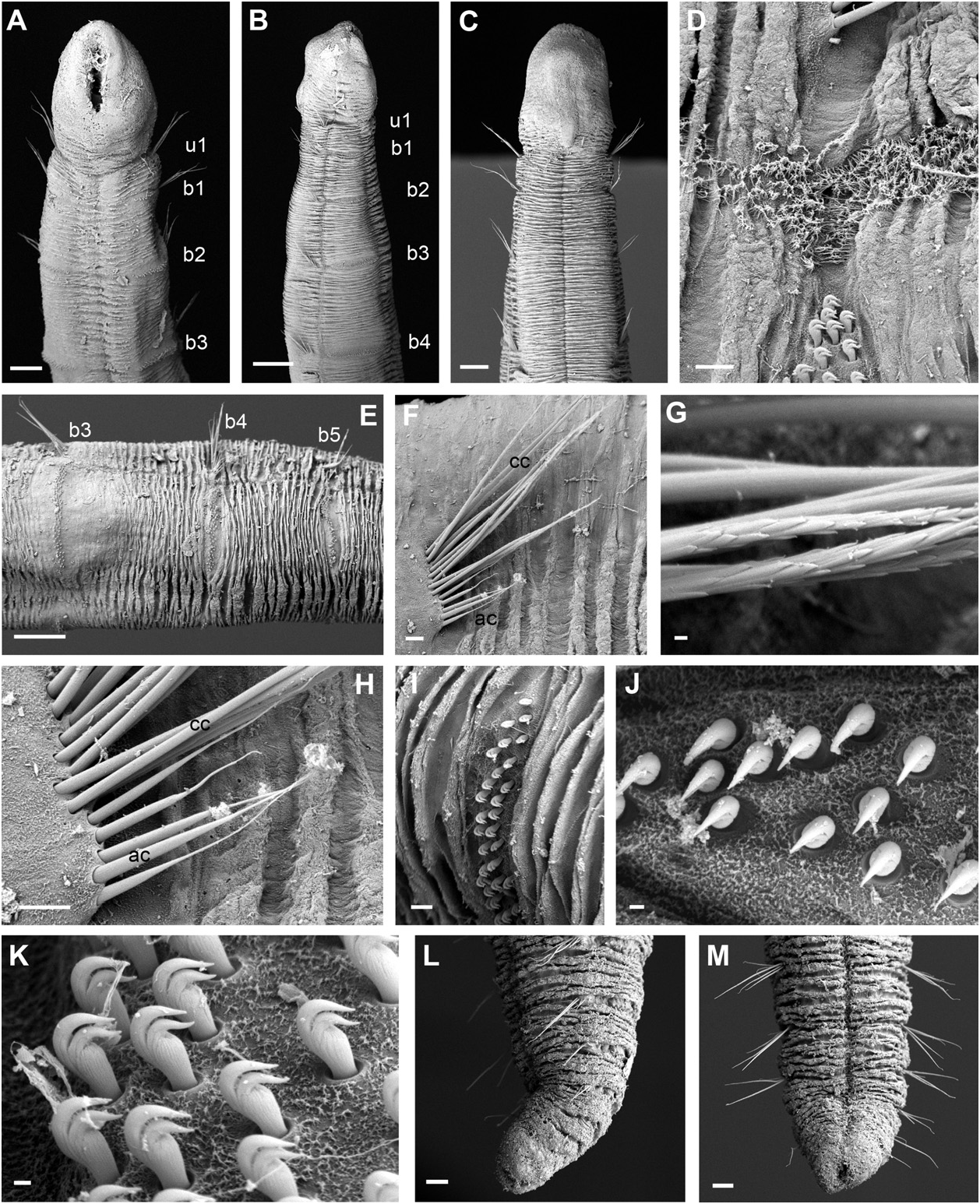
Description of holotype: Specimen measuring 7.5 mm long and 0.5 mm wide, with 26 chaetigers. Body cylindrical in cross section ( Fig. 10A View Figure 10 ) with tapering anterior and posterior ends ( Figs 10B–G View Figure 10 ; 11A–C, L, M View Figure 11 ). Epithelium wrinkled. Longitudinal ciliated grooves present along body in ventral, lateral, and dorsal sides, the dorsal being wider and more obvious ( Fig. 11C–E View Figure 11 ). Head rounded, slightly wider than the rest of body, with anterior margin slightly pointed from ventral view and distinctly set off from segmented region by constriction ( Figs 10B–D View Figure 10 ; 11A–C View Figure 11 ). Eyespots not observed. First chaetiger uniramous ( Fig. 11A, B View Figure 11 ); second and third biramous, increasing *The author described it as ‘rounded to quadrangular’.
B, biramous; nd, no data available; RLAS, relative length of anterior segments; U, uniramous. in length progressively, with RLAS = 1:2:3 ( Fig. 11A– C View Figure 11 ). Following chaetigers similar in length to posterior end, where they are progressively shorter and thinner ( Fig. 10E–G View Figure 10 ). Notopodia with capillary chaetae with proximal third smooth and distal end with scale covering ( Fig. 11G View Figure 11 ), diminishing in length ventrally within each fascicle. Acicular chaetae present from segment 5 ( Fig. 11F View Figure 11 ), about one third the length of capillaries with no ornamentation and abruptly tapered tips ( Fig. 11H View Figure 11 ). Neuropodial tori narrow and nearly rectangular, with uncini arranged in two to three irregular rows in anterior segments ( Fig. 11I, J View Figure 11 ) to three to four in middle segments ( Fig. 11K View Figure 11 ). Uncini with two teeth arranged vertically, similar in size, curved and with tips slightly pointing upwards ( Fig. 11K View Figure 11 ). Posterior end cylindrical, distally tapered, with compressed segments slightly directed dorsally ( Fig. 11L View Figure 11 ). Pygidium consisting of two low lateral lobes ( Fig. 11M View Figure 11 ). Tube with cylindrical middle section and long tapering ends, covered with sponge spicules ( Fig. 10H View Figure 10 ).
Variation: Specimens found to be between 4–8 mm in length and 0.4–0.6 mm in width with 24–35 segments, with the exception of those exhibiting anterior or posterior regeneration. The shape of the head varies amongst the specimens, being in some wider than the rest of the body but in others not obviously wider. Anterior shape also diverges amongst specimens, being pointed to rounded. Brownish lateral eyespots have been observed on the head of some specimens, but are not always present (like in the holotype), so they probably fade after preservation. Several specimens showed posterior end regenerating at different stages ( Fig. 9E View Figure 9 ).
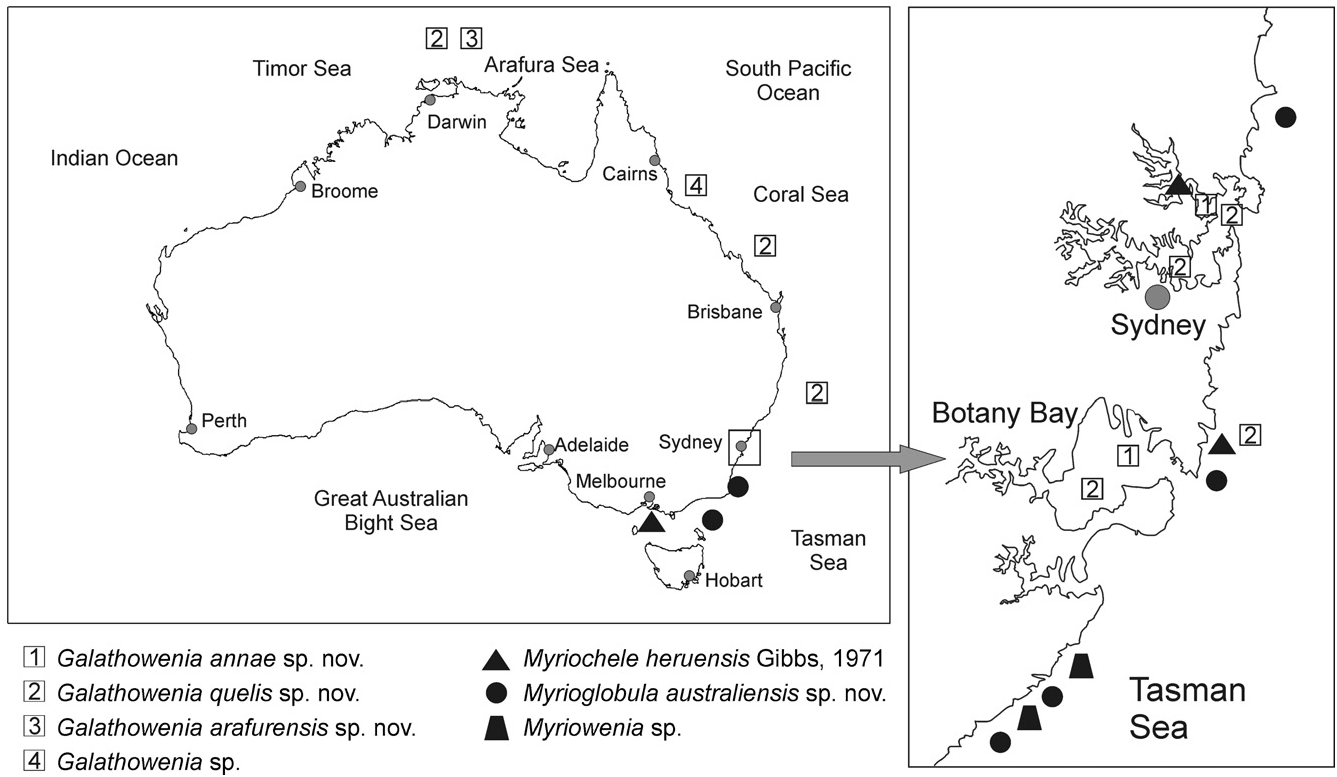
Ecological notes: Myriochele australiensis sp. nov. has been found in several localities in New South Wales, in open water environments (not estuaries) ( Fig. 14 View Figure 14 ) at around 60–80 m depth and mostly in sandy sediments. It is sympatric with Myrioc. heruensis and G. quelis sp. nov. (see Fig. 14 View Figure 14 and Appendix 2).
Etymology: The name of this species follows the tradition of many species of the former genus Myrioglobula of naming the taxon after the country in which it was discovered, in this case Australia.
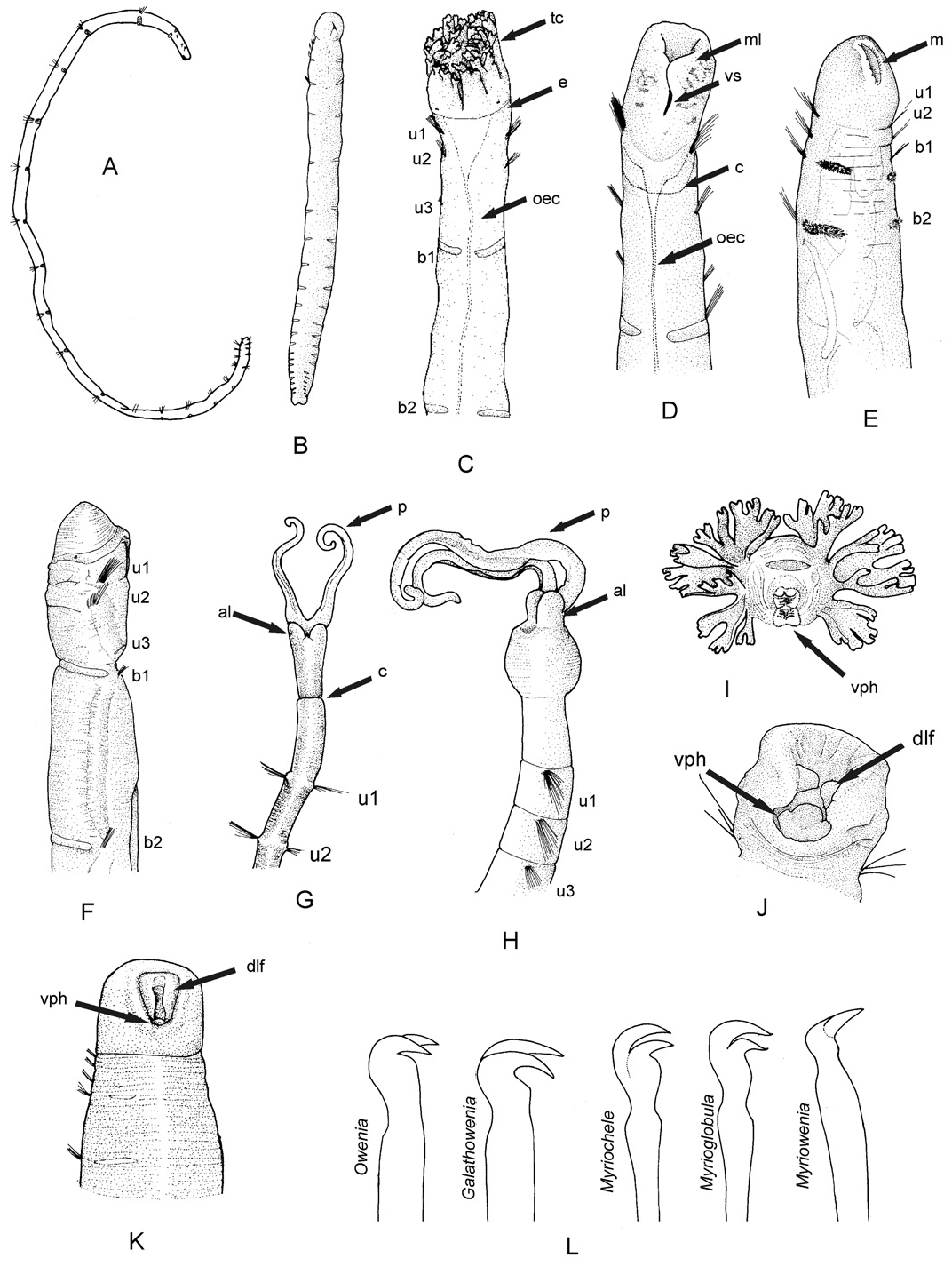
Remarks: Myriochele australiensis sp. nov. is characterized by the presence of only one anterior uniramous chaetiger, which using the traditional classification would have classified it as a species of Myrioglobula . This new species is unique amongst other species sharing this feature because it possesses peristomial eyes ( Table 5), although this feature should be used with caution as eyespots appear to fade in preserved material and could have been overlooked in the other taxa. Myriochele australiensis sp. nov. is distinguished from Myriochele antarctica ( Hartman, 1967) comb. nov. in the arrangement of the uncinal teeth, positioned in a vertical row in Myrioc. australiensis sp. nov. and other species of the genus but side by side in Myrioc. antarctica ( Table 5). Myriochele australiensis sp. nov. shares with Myriochele malmgreni ( Parapar, 2006) comb. nov., from Iceland, the shape of the pygidium with two blunt lobes; unlike Myriochele islandica ( Parapar, 2003a) comb. nov., which has a tetra-lobed pygidium and Myriochele japonica ( Imajima & Morita, 1987) comb. nov., with a pygidium consisting of two large lobes and five papillae. However, Myrioc. australiensis sp. nov. and Myrioc. malmgreni comb. nov. differ in the relative length of anterior segments (1:2: 3 in the former and 1:1.5: 5 in the latter) and in the appearance of the anterior-most acicular chaetae on notopodia, being in the fifth segment in Myrioc. australiensis sp. nov. and in the third in Myrioc. malmgreni comb. nov.
The uncini of Myrioc. australiensis sp. nov. ( Fig. 11K View Figure 11 ) show a characteristic ‘S’ shape with a prominent subrostral process and the tip of the teeth pointing upwards slightly ( Fig. 15L View Figure 15 ). These features were previously observed in three of the four species hitherto described in the genus Myrioglobula : Myrioglobula japonica ( Imajima & Morita, 1987; Figs 9F View Figure 9 , 10F View Figure 10 ), Myrioglobula islandica ( Parapar, 2003a; Fig. 8C View Figure 8 ), and Myrioglobula malmgreni ( Parapar, 2006; Fig. 11G View Figure 11 ) but not in the Myrioglobula type species: Myrioglobula antarctica .
Myriochele australiensis View in CoL sp. nov. resembles Myriochele minor Caullery, 1944 View in CoL , recorded from the Flores Island, Indonesia, in the shape of the head, elongated and with a rounded anterior end, and in the shape of the mouth. However, they differ in thoracic formula, U: B: B in Myrioc. australiensis View in CoL sp. nov. and U: U: U in Myrioc. minor View in CoL ; in the shape of the pygidium, bilobed in the new species and with three lobes (to dorsal and one ventral) in Myrioc. minor View in CoL ; and in the relative length of the anterior segments, RLAS = 1:2:3:3:4: 3 in Myrioc. australiensis View in CoL sp. nov. and RLAS = 1:1:1:1: 4 in Myrioc. minor View in CoL .


| AM |
Australian Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Myriochele australiensis
| Capa, Maria, Parapar, Julio & Hutchings, Pat 2012 |
Myriochele australiensis
| Capa & Parapar & Hutchings 2012 |
australiensis
| Capa & Parapar & Hutchings 2012 |
australiensis
| Capa & Parapar & Hutchings 2012 |
Myriochele minor
| Caullery 1944 |
minor
| Caullery 1944 |
minor
| Caullery 1944 |
minor
| Caullery 1944 |