
Myriochele, MALMGREN, 1867
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2012.00850.x |
|
persistent identifier |
https://treatment.plazi.org/id/03AE8729-AB0E-D442-FCE2-FA5CA59DF947 |
|
treatment provided by |
Marcus |
|
scientific name |
Myriochele |
| status |
|
GENUS MYRIOCHELE MALMGREN, 1867 View in CoL , EMENDED
Myriochele Malmgren, 1867 View in CoL ; Blake, 2000; Parapar, 2006.
Myrioglobula Hartman, 1967 View in CoL .
Emended diagnosis: Body short and thick, tapering at both ends (cigar-like). Head lacking appendages, with anterior end rounded. Mouth terminal or slightly elongated ventrally but without a ventral slit and developed lips on the sides. Anterior one to three segments uniramous, subsequent ones biramous. Notopodia with capillary and acicular chaetae (latter type often absent in anterior-most chaetigers). Neuropodial tori with uncini bearing two similar-sized teeth generally vertically arranged, but obliquely in some species.
Type species: Myriochele heeri Malmgren, 1867 , by original designation.
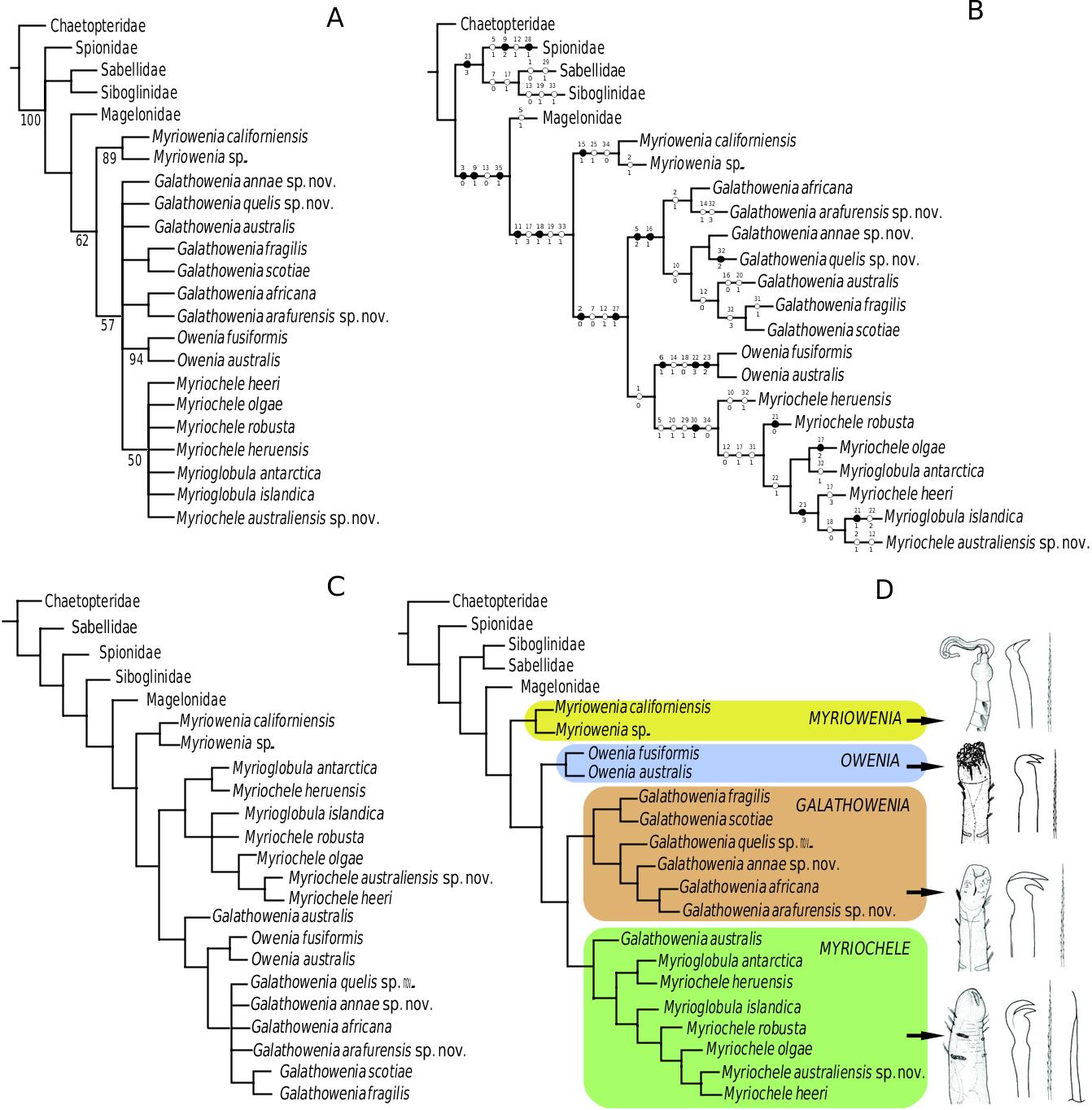
Remarks: Our phylogenetic analyses recover Myrioglobula species scattered within those traditionally assigned to Myriochele ( Fig. 1A–D View Figure 1 , unlike Sene-Silva, 2002). Although the support value for this clade is not strong ( Fig. 1A View Figure 1 ), the morphological features maintaining the two genera cannot be justified. Differences between Myriochele and Myrioglobula were traditionally based on the number of uniramous segments ( Hartman, 1967), with Myrioglobula characterized by having one (TF = U: B: B) and Myriochele two or three (TF = U: U: B and U: U: U). However, this feature has been demonstrated not to be informative because it varies within Myriochele (two or three segments), and there are species, such as Myriochele robusta Parapar, 2003c , which show a different pattern with a biramous second segment between the uniramous first and third (TF = 1:0–1:1–1:0 sensu Parapar, 2003c and U: B: U in this study). Features shared by the two groups are the shape of the body and head, presence of acicular chaetae, and arrangement of uncinal teeth; strong enough, according to the phylogenetic hypothesis presented here, to consider Myrioglobula as a junior synonym of Myriochele . We also propose here to include G. australis within the genus Myriochele , a species with acicular chaetae on parapodium and a poorly defined anteriorly truncated head and ventral slit ( Parapar, 2003b; see Fig. 1D View Figure 1 ), regardless of the possible variation of the arrangement of uncinal teeth in the genus Galathowenia . Myriochele is the most morphologically diverse genus in the family Oweniidae with over 21 species known ( Villalobos-Guerrero, 2009).
MYRIOCHELE HERUENSIS GIBBS, 1971 View in CoL
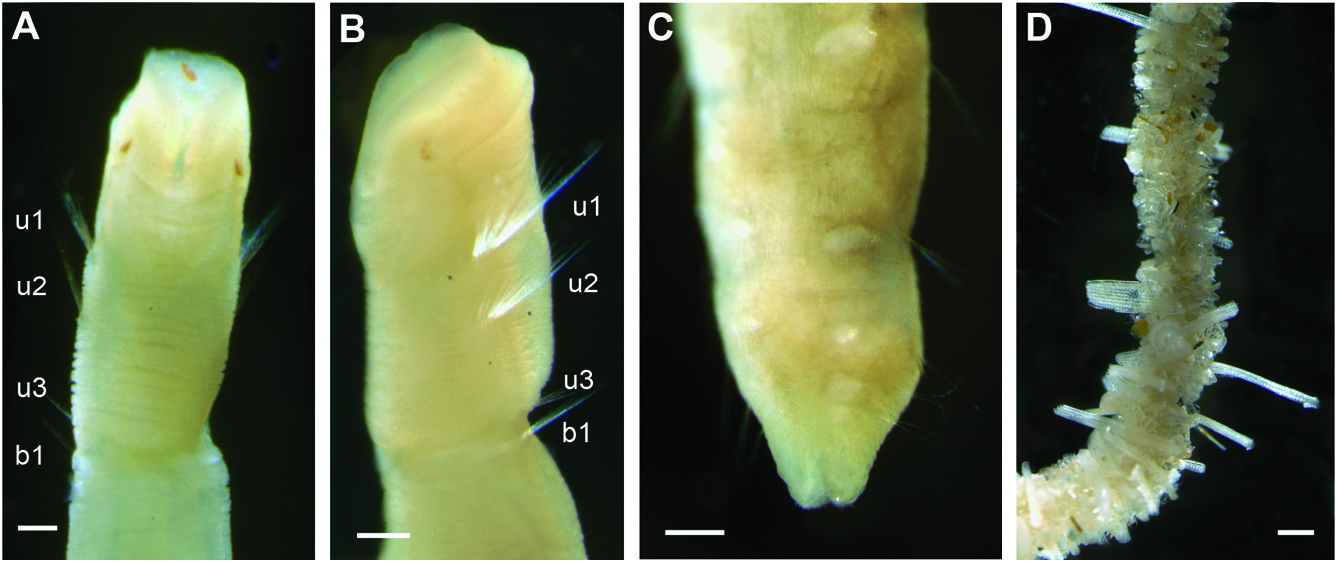
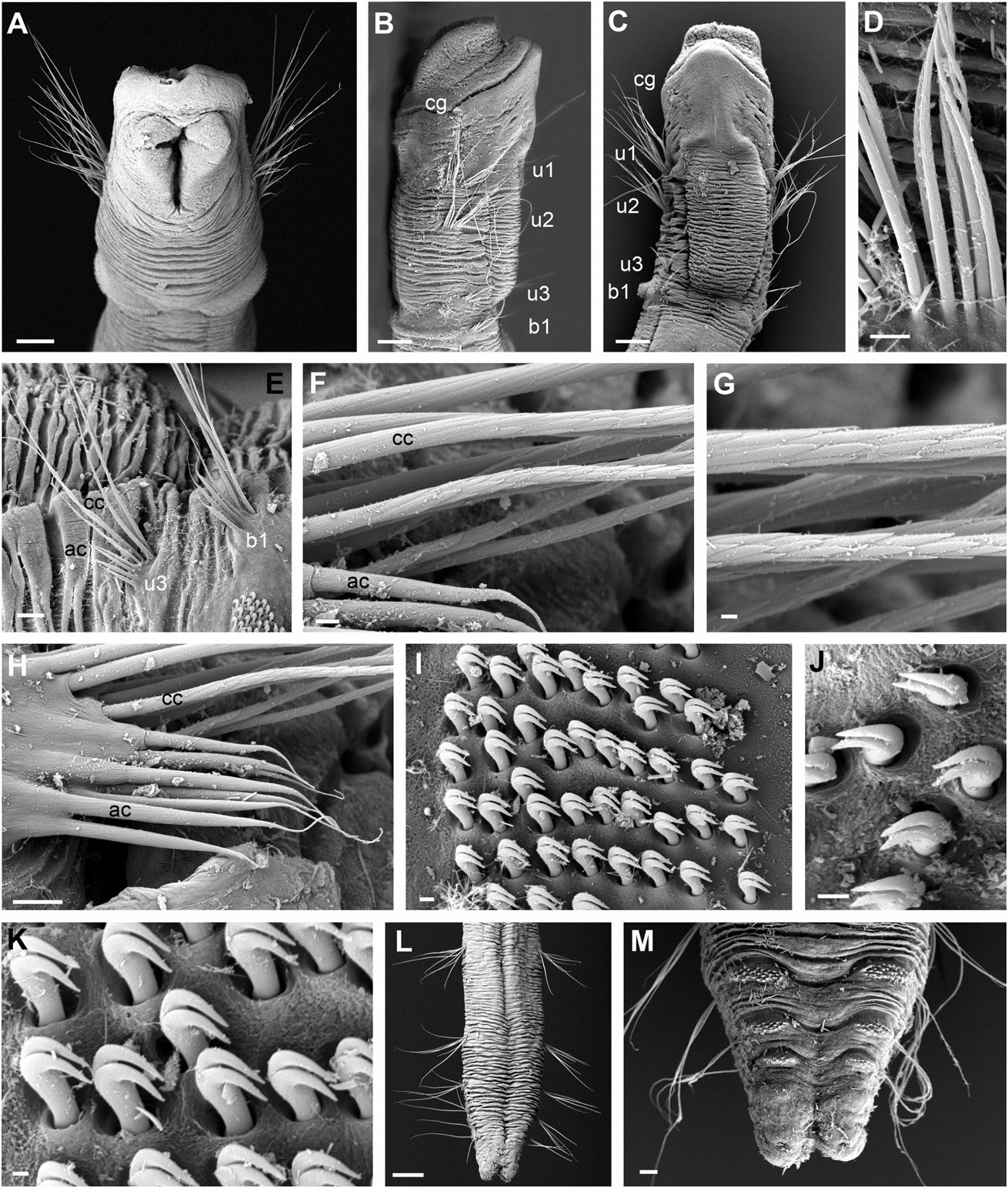
FIGURES 7 View Figure 7 , 8 View Figure 8 , 9A–D, TABLES View Figure 9 1, 4
Myriochele heruensis Gibbs, 1971: 190–191 View in CoL , figure 15B–G.
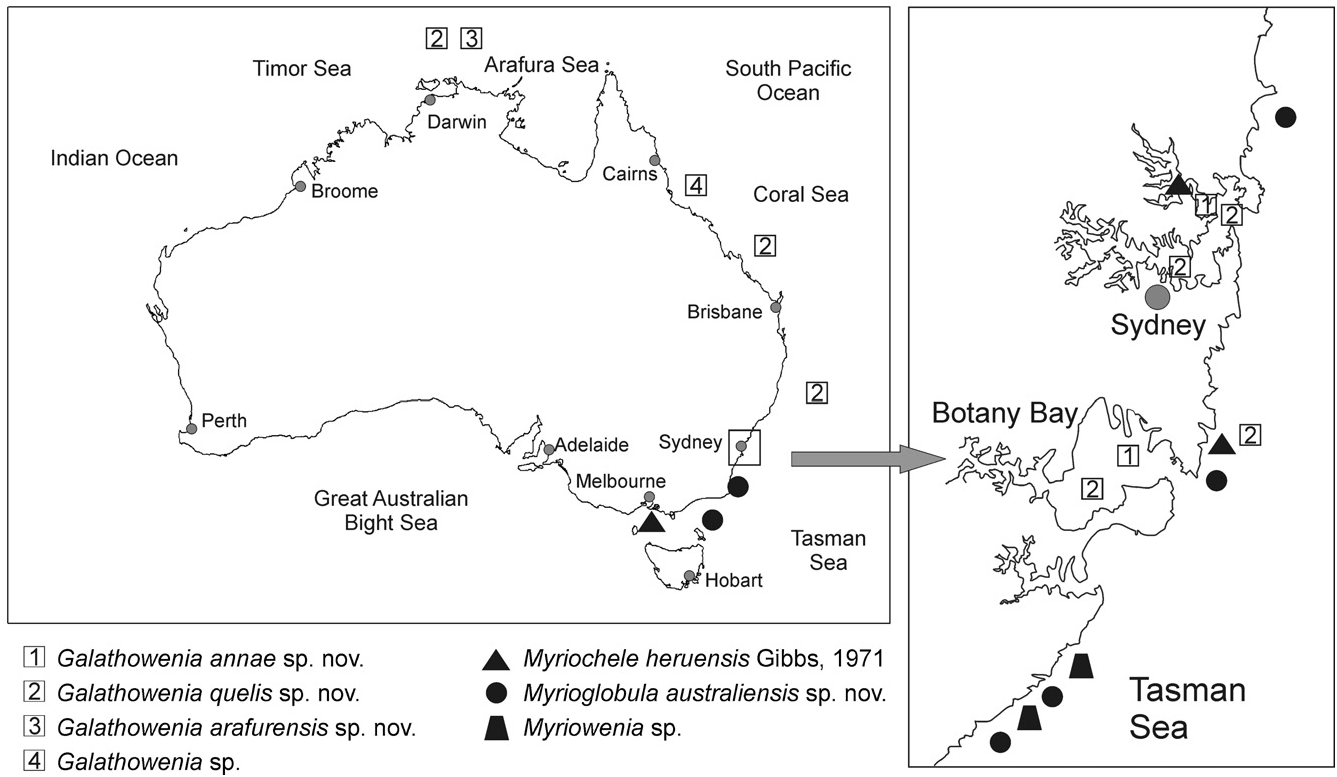
Material examined (Appendix 1): Victoria, Port Phillip Bay (828 specs) ; New South Wales (264 specs): Malabar, Pittwater , between 1 and 60 m, in sand and muddy sand .
Description of Australian specimens: Body long, cylindrical, narrowing at posterior end, measuring 4–20 mm long, 0.2–0.5 mm wide, consisting of 12–18 segments. Epithelium wrinkled. Head similar in width to rest of body and swollen dorsally resembling a ‘humpback’ and divided from the segmented region by a deep groove, oblique from side view ( Figs 7A, B View Figure 7 , 8A–C View Figure 8 ). Anterior shape of prostomium varies amongst specimens, appearing as rounded or truncated depending on the shape of the mouth opening and the development of the ‘humpback’, whose size changes between individuals. Pair of lateral red eyespots located on ventrolateral side, just below groove ( Fig. 7A, B View Figure 7 ). Mouth anterior-ventral ( Figs 7A View Figure 7 , 8A View Figure 8 ), with two large lateral lips ( Figs 7A, B View Figure 7 , 8A, B View Figure 8 ). First three segments uniramous, with only notochaetae, no segmental grooves between them. First chaetiger about half length of second and twice that of the third one ( RLUS = 2:4:1). Oesophageal commissures evident as a white ventral longitudinal band. First two chaetigers with long capillary notochaetae ( Figs 7A, B View Figure 7 , 8A–C View Figure 8 ), compared to subsequent ones. Acicular chaetae arranged in ventral-most part of fascicle, from segment 3 ( Fig. 8E, F, H View Figure 8 ). Capillary chaetae with proximal part smooth or slightly striated ( Fig. 8D View Figure 8 ) and mid and distal end with sculptures resembling scales ( Fig. 8F, G View Figure 8 ); acicular chaetae smooth ( Fig. 8H View Figure 8 ). Chaetigers biramous from segment 4. First biramous chaetiger at least 1.5 times as long as uniramous region; second and third biramous segments (chaetigers 5 and 6) twice the length of first biramous; fourth biramous chaetiger the longest, about three times longer than first biramous chaetiger ( RLFBS = 1:2:2:3). Chaetae of first biramous segment (chaetiger 4) located just posterior to notochaetae of third chaetiger ( Fig. 8E View Figure 8 ). Biramous segments with capillary and acicular chaetae on notopodia, similar to those described in anterior segments. Neuropodial tori with about ten rows of teeth on anterior segments ( Fig. 8I View Figure 8 ). Uncini bidentate, with curved teeth and tips pointing upwards, obliquely *Only in some populations.
†Although author’s drawings and formula of relative length do not correspond with this observation (see Nilsen & Holthe, 1985: fig. 5).
‡Although the types were collected in the Solomon Islands , the scoring of this table has been completed with additional material from Australia .
nd, no data available; RLFBS, relative length of first biramous segments; RLUS, relative length of uniramous segments.
arranged ( Fig. 8J, K View Figure 8 ). Posterior chaetigers becoming shorter and narrower, ending in a narrow pygidium provided with two lateral lobes (( Figs 7C View Figure 7 , 8L, M View Figure 8 ). Most specimens are colourless or pale yellow ( Fig. 7A–C View Figure 7 ) but others have a row of pigment along the anterior margin of the head. Similarly, in some specimens, the eyespots seem to have faded with preservation. Several specimens showed anterior or posterior ends regenerating at different stages ( Fig. 9A–D View Figure 9 ), with anterior regeneration being the most common. Tube encrusted with sea urchin spines and sand grains in no regular pattern ( Fig. 7D View Figure 7 ).
Ecological notes: Myriochele heruensis was previously reported on sandy bottoms at 16 m depth ( Gibbs, 1971). Our specimens were found in sheltered bays and estuaries of New South Wales and Victoria and in some deep water exposed environments ( Fig. 14 View Figure 14 ), on sand and muddy sand substrates from 15 to 85 m depth .
Distribution: The species was originally described from the Marovo Lagoon (Vangunu Island) in the New Georgia Group of the Solomon Islands ( Gibbs, 1971). This is the only report of the species since the original description.
Remarks: We have assigned these specimens to My. heruensis as both populations share several morphological features considered as diagnostic. This species is characterized by the presence of a dorsal swollen area on the head, an anterior groove located across the dorsal surface of the prostomium and with an anterior margin of the head that varies amongst specimens. This head plasticity had already been reported by Gibbs (1971) who highlighted that the anterior groove is shallow when the prostomium is protracted and appears as a fold when it is contracted. Another two diagnostic features are the presence of a distinctive oblique cephalic groove (dorsolateral sulcus, according to Nilsen & Holthe, 1985) and an enlarged segment 7. Gibbs (1971) does not report the presence of acicular chaetae; in our opinion they probably were unnoticed by the author because other characters – body size, RLUS, shape of uncini and pygidium, and especially the head characteristics (prostomial groove and dorsal humpback) – are fully consistent with our material.
Other species of Myriochele sharing a similar head shape are Myriochele danielsseni Nilsen & Holthe, 1985 , (European Atlantic and? Japan) and Myriochele striolata Blake, 2000 , (East Pacific) which also share with the new species the large number of transverse wrinkles along the body (numerous transverse ventral thoracic furrows, according to Blake, 2000). Myriochele heruensis differs from the original description of Myrioc. danielsseni in the relative number and the length of anterior biramous segments (see Table 4). Segment 7 (fourth biramous chaetiger) is the longest in the new species ( RLFBS = 1:2:2:3) and segments 5–7 (second to fourth biramous chaetiger) are the longest in Myrioc. danielsseni ( RLFBS = 1:1.5:1.5:1.5). The specimens identified as Myrioc. danielsseni from Japan ( Imajima & Morita, 1987) and proposed here to be conspecific with Myrioc. striolata ( Blake, 2000) differ from the original description of both species and from Myrioc. heruensis in that feature, as segments 6–8 are the most elongated (see Table 4 for comparisons amongst the species). Myriochele heruensis is distinguished from Myrioc. striolata by the overall number of body segments, with more than ten present in the new species and always with fewer than ten present in Myrioc. striolata ( Blake, 2000) , and in the position of the most elongated segment, being the fourth biramous chaetiger (segment 7) in Myrioc. heruensis and the second biramous chaetiger (segment 5) in Myrioc. striolata . Moreover, Myrioc. heruensis possesses acicular chaetae on notopodia from segment 3 whereas this type of chaetae is apparently absent in Myrioc. striolata ( Blake, 2000) . Myriochele heruensis and the Japanese population of Myrioc. danielsseni are the only species described as having pigment bands on the head, whereas the two other taxa have been described as lacking colour pigment ( Nilsen & Holthe, 1985; Blake, 2000). These two species are also distinguished by the other features mentioned above (see also Table 4). Myriochele danielsseni, Myrioc. striolata , and Myrioc. heruensis form a well-characterized group of species sharing several morphological features, as stated above. Specimens described and illustrated by Gibbs (1972) as Myriochele sp. , from the Cook Islands resemble those described herein as Myrioc. heruensis . However, some features cannot be compared because the three specimens of Myriochele sp. were incomplete and there is no mention of the acicular chaetae. It would be interesting to assess the relationships of this group of species with other species of Myriochele in a phylogenetic framework.
The finding of an individual split into two parts inside the tube, with each part in the process of regenerating the missing half, could be an indication of reproduction by schizotomy as described for Myrioc. danielsseni ( Aguirrezabalaga, Gil & Viéitez, 2000) . However, we cannot confirm this type of reproduction in our material.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Myriochele
| Capa, Maria, Parapar, Julio & Hutchings, Pat 2012 |
Myriochele heruensis Gibbs, 1971: 190–191
| Gibbs PE 1971: 191 |