
Galathowenia annae, Capa & Parapar & Hutchings, 2012
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2012.00850.x |
|
DOI |
https://doi.org/10.5281/zenodo.10544620 |
|
persistent identifier |
https://treatment.plazi.org/id/03AE8729-AB1B-D45D-FEAE-FC9BA753F9EE |
|
treatment provided by |
Marcus |
|
scientific name |
Galathowenia annae |
| status |
sp. nov. |
GALATHOWENIA ANNAE View in CoL SP. NOV.
FIGURES 2A–D View Figure 2 , 3 View Figure 3 , TABLES 1, 2
Holotype: New South Wales, Botany Bay , east end of runway 34 R, AM W37222, 33°58′S, 151°12′E, sandy mud, 16 m, 15.ix.2004. GoogleMaps
Paratypes: New South Wales, Botany Bay , east end of runway 34 R, AM W37223, same sample [five specimens (specs)] ; AM W37208, same sample (one spec. on SEM pin) ; AM W37224, 33°58′31″S, 151°11′49″E, mud, 16 m, 1.xii.2004 (four specs) and GoogleMaps AM. W37064, same sample (one spec. on SEM pin) ; AM W37225, 33°57′37″S, 151°11′15″E, mud, 7.4 m, 1.xii.2004 (15 specs) GoogleMaps ; AM W37226, 33°58′38″S, 151°11′45″E, mud, 18.6 m, 1.xii.2004 (one spec.) GoogleMaps ; AM W37227, 33°57′43″S, 151°11′18″E, mud, 7.6 m, 1.xii.2004 (four specs) GoogleMaps ; AM W37228, 33°58′40″S, 151°11′49″E, sandy mud, 18.2 m, 15.ix.2004 (two specs) GoogleMaps .
Other material examined (Appendix 1): New South Wales, Botany Bay (34 specs), Pittwater (one spec.) .
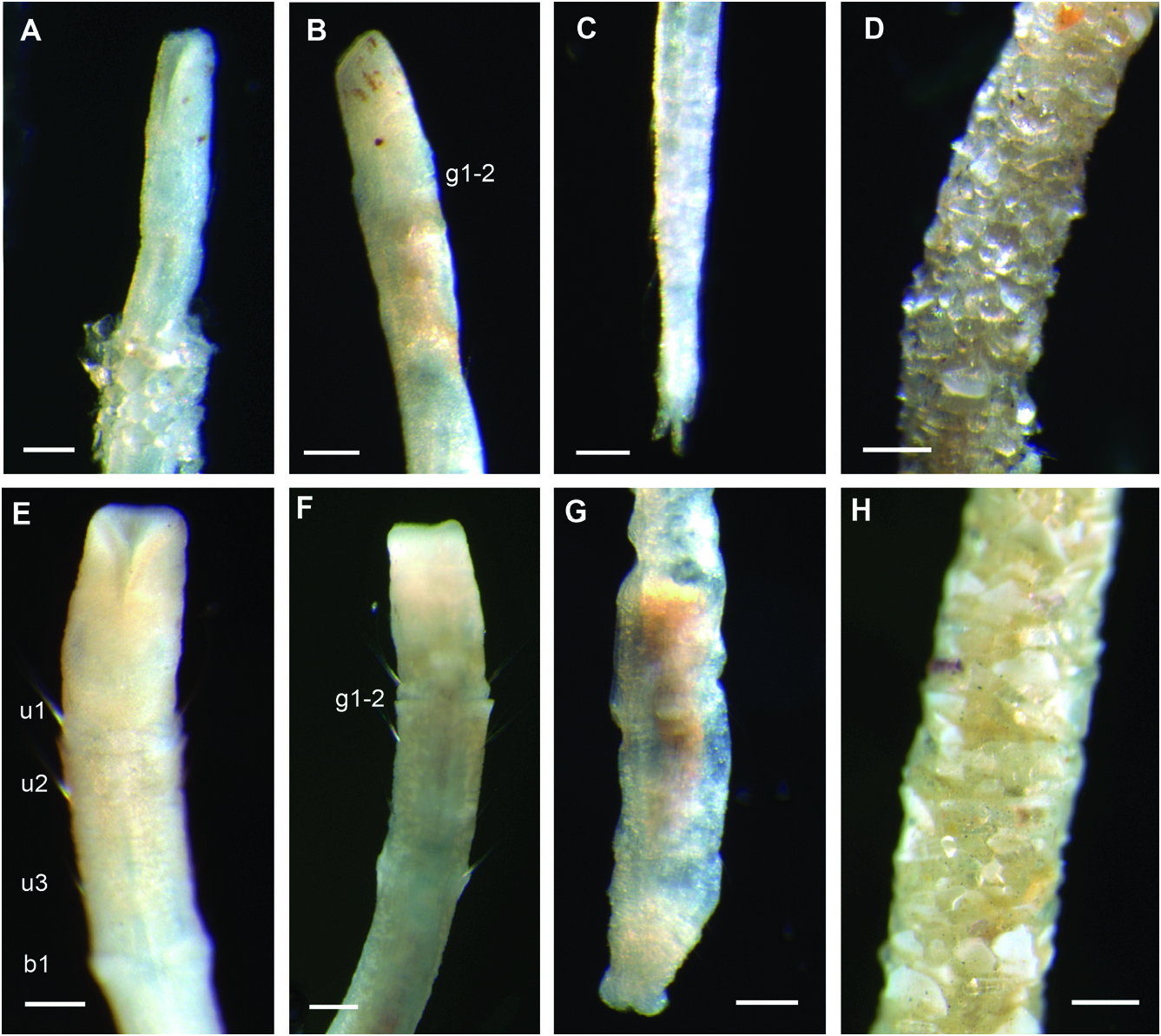
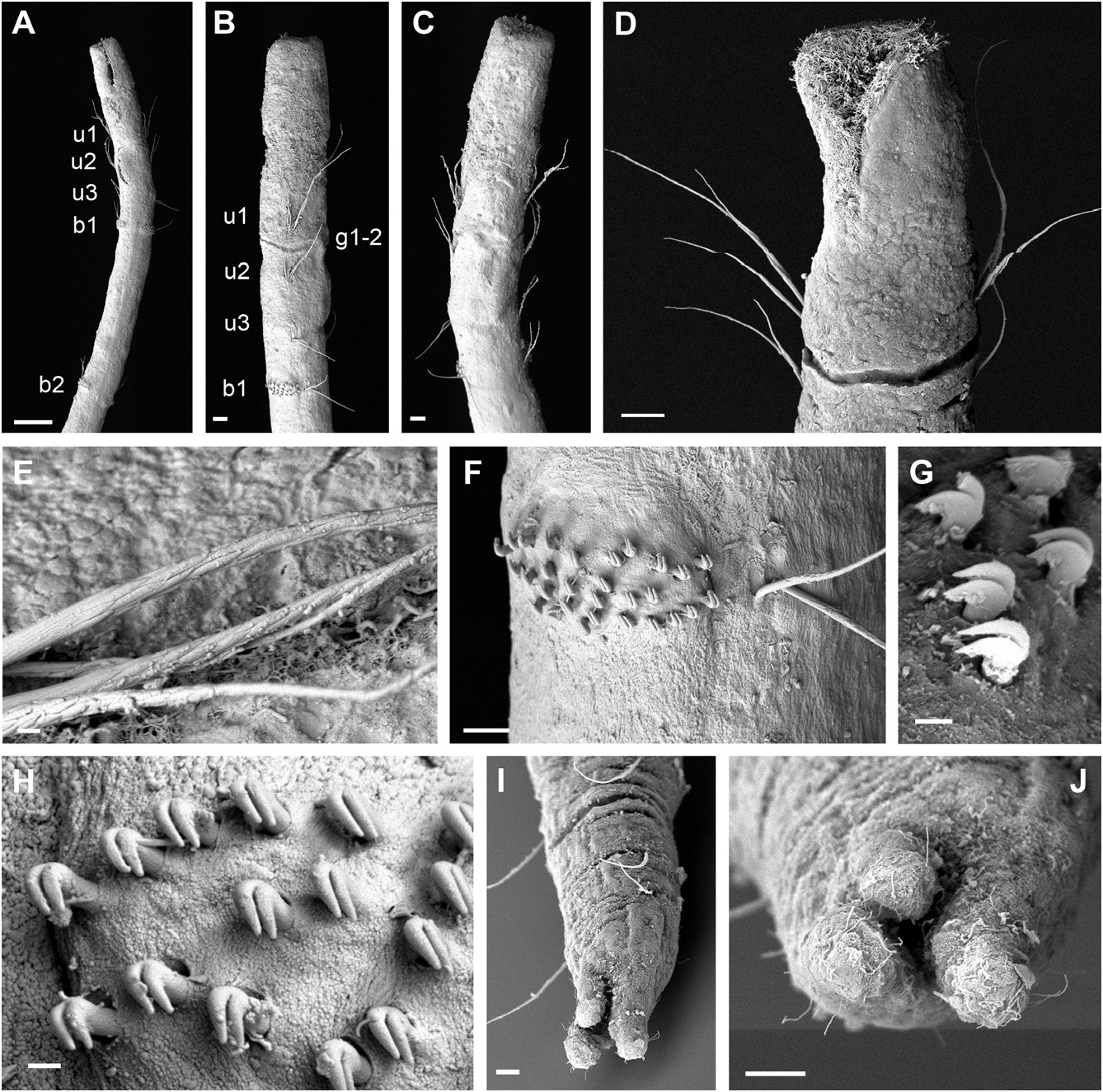
Description of holotype: Slender, thread-like body, cylindrical in cross section, 10.2 mm long, 0.1 mm wide, with 17 chaetigers. Head elongated with anterior end truncated; terminal ciliated mouth opening, extending midventrally as elongated slit ( Figs 2A, B View Figure 2 , 3A–D View Figure 3 ), with a pair of ventrolateral brownish eyespots ( Fig. 2A, B View Figure 2 ). Division of head and first segment inconspicuous, no groove or variation in width apparent. Anterior three segments with uniramous parapodia, each with notochaetae only ( Fig. 3A, B View Figure 3 ). Second segment twice as long as first and third; RLUS = 1:2:1 ( Fig. 3B View Figure 3 ). Deep groove between first and second chaetigers, on ventral and lateral sides ( Fig. 3B, D View Figure 3 ). Oesophageal commissures evident as a white ventral longitudinal band with a Y shape in anterior segments. Fourteen biramous chaetigers. Anterior biramous chaetigers five to more than ten times longer than wide (e.g. Fig. 3A View Figure 3 ), becoming shorter and compacted in far posterior segments. Chaetiger 6 longest. Capillaries decreasing in size ventrally on notopodia with proximal part smooth or slightly striated and mid and distal end covered with scales ( Fig. 3E View Figure 3 ). Posterior chaetigers with fewer but longer chaetae. Neurochaetae present from chaetiger 4, in long and broad ventral tori; tori of posterior segments much shorter. Acicular chaetae absent. Uncini arranged in about four irregular transverse rows in anterior segments ( Fig. 3F View Figure 3 ); each uncinus provided with two convex teeth arranged in an oblique row ( Fig. 3G, H View Figure 3 ). Anus terminal with three prominent digitiform lobes, dorsal one slightly smaller than lateral ones ( Fig. 3I, J View Figure 3 ). Tube three to four times longer than animal, with thin secreted layer encrusted with sand grains ( Fig. 2D View Figure 2 ). Colour in alcohol brownish or pale yellow, head with short brownish red pigmented areas ( Fig. 2B View Figure 2 ) and some slightly pigmented bands in anterior four segments.
Variation: All specimens examined are cylindrical in shape and very thin, not exceeding 0.1 mm in width. Length and number of segments vary between 5–11 mm and from 14 to 17 segments. Most specimens were incomplete or difficult to extract from their tubes so variation could be greater than reported herein. The colour pattern varies amongst the material examined. Some specimens have a fully pigmented brownish head and first segments whereas in others the body is almost colourless or presents only few scattered brownish pigments spots. Eyespots have faded in some specimens, probably because of the length of time in alcohol. In most specimens pygidial lobes are of similar size, but the dorsal lobe has been found to vary from being shorter or longer than lateral ones.
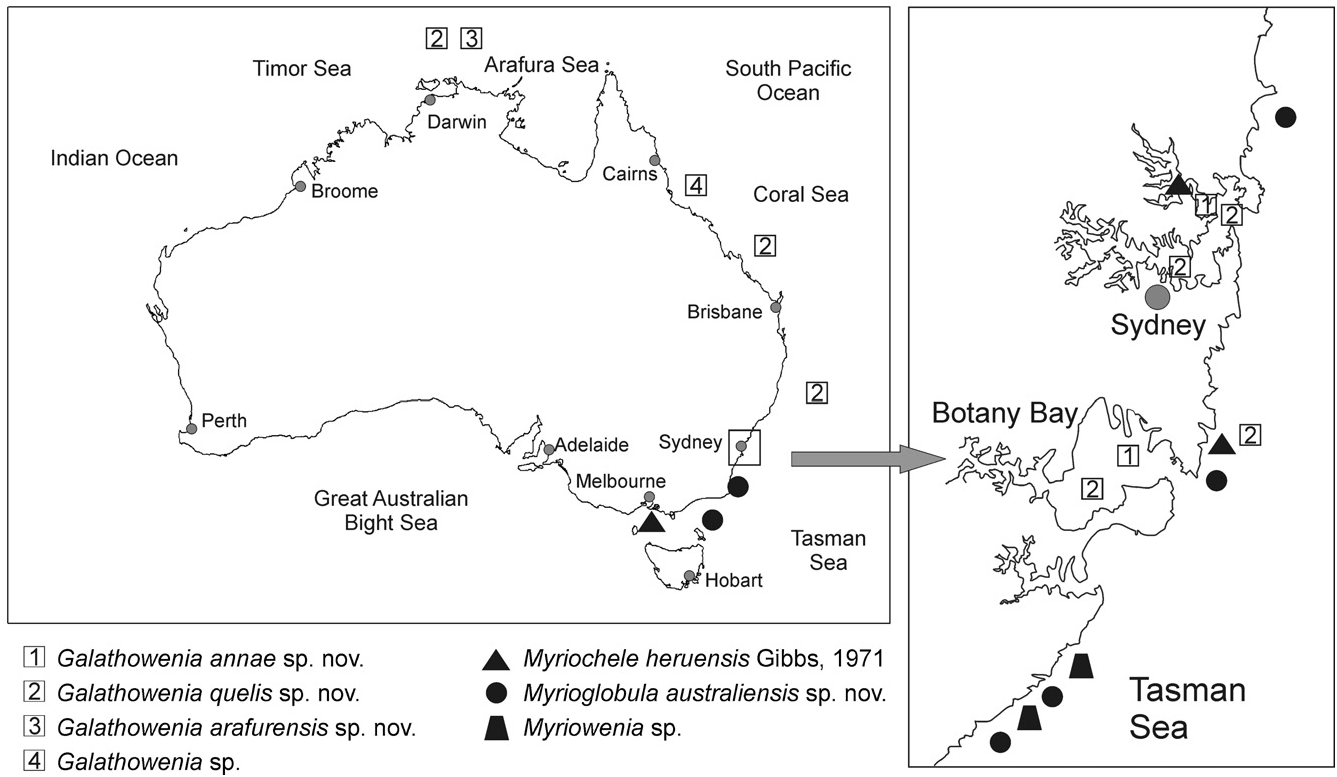
Ecological notes: Galathowenia annae sp. nov. lives in estuaries of the Sydney region ( Fig. 14 View Figure 14 ), inhabiting muddy and sandy sediments between 13 and 18 m in depth.
Etymology: This species is dedicated to Anna Murray (AM, Sydney) for providing valuable preliminary notes on the identification of Australian oweniids.
Remarks: Species of Galathowenia are characterized by a combination of only a few relevant features, a reason why some specimens collected around the globe have been attributed to a few ‘cosmopolitan species’. This is the case for Galathowenia oculata ( Zachs, 1923) , originally described from the White Sea ( Zachs, 1923) and reported in several localities from boreal to American Pacific regions, and depths ranging from 12 to 2500 m (e.g. Nilsen & Holthe, 1985; Imajima & Morita, 1987; Blake, 2000; Parapar, 2003a). Some of the diagnostic features present in G. oculata , such as the presence of eyespots, short anterior uniramous segments, long biramous segments (particularly the fifth), and pygidium provided with two or three blunt lobes are commonly present in specimens across the broad range of its geographical distribution. However, specimens from different biogeographical areas show several differences that some authors have recognized as interpopulation variation, but that could be also interpreted as specific differences. Accordingly, we interpret G. oculata as a species complex to which G. annae sp. nov., belongs, but can be distinguished based on a range of characters ( Table 2). Galathowenia annae sp. nov. is characterized by its thread-like body, being one of the thinnest species described in Galathowenia , and the presence of a pygidium with three prominent digitiform lobes, two of which are ventrolateral and the third dorsal. These lobes have only been described in G. oculata from the western Mediterranean ( Martín, 1989), which could represent another undescribed species based on this and other features ( Blake, 2000; Parapar, 2003a). After the examination of two specimens from the Mediterranean we verified the similarities between these and G. annae sp. nov. They share the presence of eyes, the pigmentation in the anterior end, the relative size of the three first *, type species of each genus.
Multistate characters: A = 0 + 1.
The generic names used in this table correspond to the classification of the species previous to the present study. *Smaller specimens are probably juveniles.
†From the study of two specimens.
‡Feature not described, information obtained from drawings of photographs.
§From Blake & Dean (1973).
¶In Icelandic populations ( Parapar, 2003a).
nd, no data available; RLUS, relative length of uniramous segments.
segments (RLUS = 1:2:1), and the presence of three pygidial cirri, one arranged dorsally and the other two lateroventrally. However, in the Mediterranean specimens of G. oculata the pygidial cirri are similar in size, the deep groove between first and second chaetiger is absent, and the longest segment is the fifth instead of the sixth (see Table 2). These differences, if confirmed in a greater number of specimens from the Mediterranean, could establish the limits between these two species and the rest of the G. oculata complex. Nilsen & Holthe (1985) described some populations of G. oculata as presenting a small dorsal lobe in the pygidium but according to the drawings ( Nilsen & Holthe, 1985: fig. 7C), it is very small compared to the one found in G. annae sp. nov. Some populations of G. oculata from northern Europe were also described with a dorsal pigmented band on the head ( Nilsen & Holthe, 1985), different from the irregular and inconstant pattern observed in the specimens of G. annae sp. nov. studied herein. The specimens of G. oculata described from Japan ( Imajima & Morita, 1987) share the colour pattern with G. annae sp. nov., but they are distinguished by the shape of the pygidium, with two lobes in the Japanese specimens and three in the Australian ones. Galathowenia annae sp. nov. presents a deep groove between the first and the second chaetiger in the specimens examined and this could be related to regeneration processes, as indicated by Parapar (2003a) for Galathowenia fragilis ( Nilsen & Holthe, 1985) . This feature has not been described in any of the hitherto-published descriptions of G. oculata (see Parapar, 2003a), although one groove between the head and the first segment was illustrated for this species based on specimens from California, USA ( Blake, 2000: fig. 5.1A) and two grooves indicated in specimens from Japan ( Imajima & Morita, 1987: fig. 6A–C). One of the specimens collected on Makassar, Sulawesi ( Indonesia) by Caullery (1944: fig. 42C) and considered as a variety of Myriochele eurystoma Caullery, 1944 , resembles G. annae sp. nov., in the shape of the head and in the colour pattern. Unfortunately, the description of this specimen is not complete and the material was not available for a detailed comparison.
| R |
Departamento de Geologia, Universidad de Chile |
| AM |
Australian Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |