
OWENIIDAE, RIOJA, 1917
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2012.00850.x |
|
persistent identifier |
https://treatment.plazi.org/id/03AE8729-AB3E-D476-FCC2-FBFEA294FE1F |
|
treatment provided by |
Marcus |
|
scientific name |
OWENIIDAE |
| status |
|
OWENIIDAE View in CoL RELATIONSHIPS AND CHARACTER
TRANSFORMATION
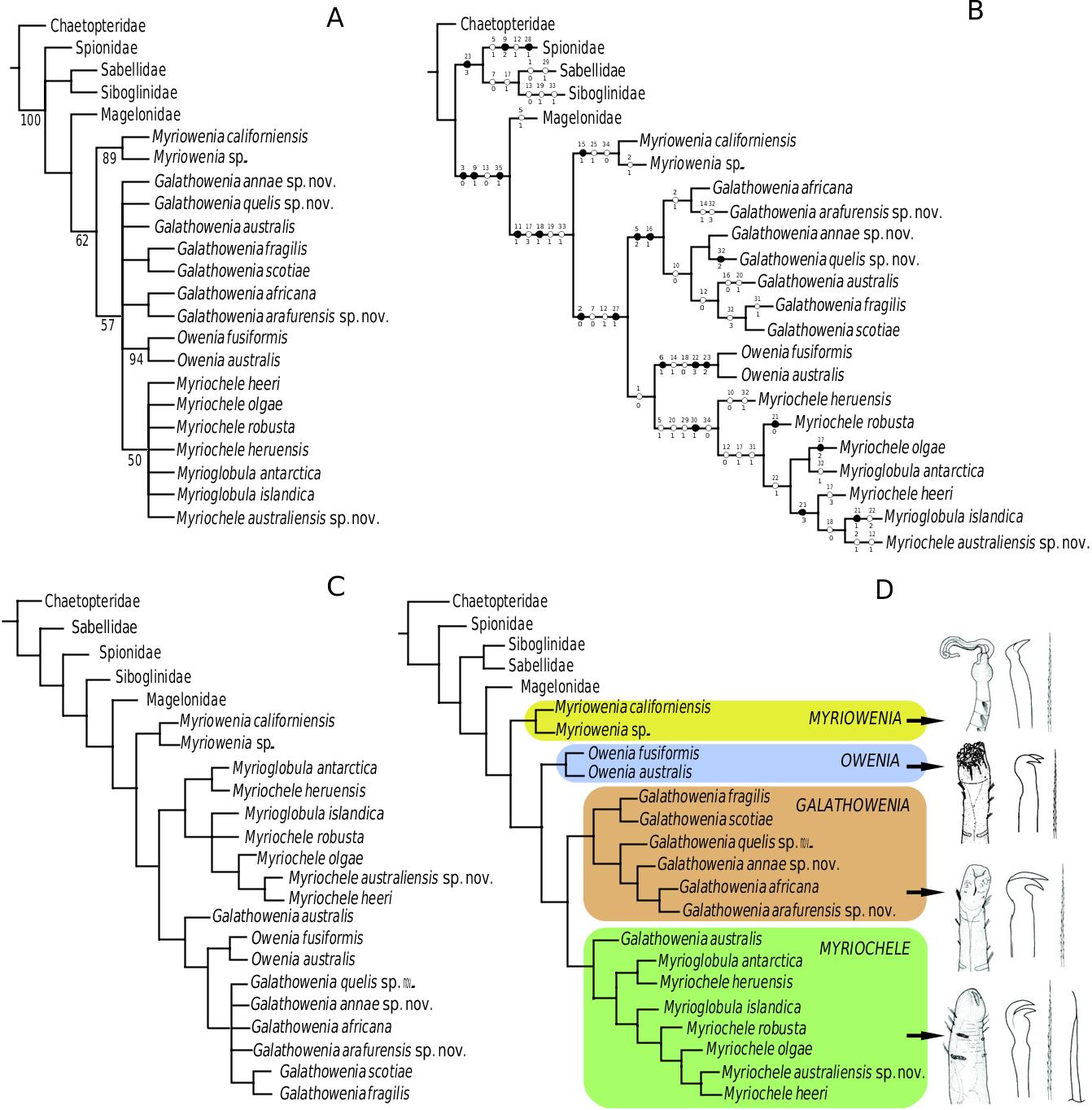
Our results endorse the monophyly of the Oweniidae (as did Sene-Silva, 2002) and four of the traditional oweniid genera if some synonymizations and nomenclatural changes are carried out. The monophyly of Owenia and Myriowenia has never been questioned owing to the presence of their characteristic head appendages (tentacular crown and grooved palps, respectively). However, the position of these groups differs in the literature. The hypothesis proposed by Sene-Silva (2002) recovered Myriowenia as the basal oweniid and Owenia as the most derived. Other authors (e.g. Nilsen & Holthe, 1985) also suggested this hypothesis based on the arrangement of uncinal teeth, with vertically arranged teeth in Myriowenia as the plesiomorphic condition, also found in other polychaetes, and side by side in Owenia as the apomorphic condition. In our hypothesis the position of Owenia depends on methodological procedures and although its position in the nonweighted analyses is uncertain, low concavity values topologies support the hypothesis of the apomorphic condition of the uncini with side by side teeth ( Fig. 1C View Figure 1 ). However, the results with higher values of k imply that the oblique arrangement of uncinal teeth is the derived condition ( Fig. 1D View Figure 1 ).
The genus Galathowenia was defined by the shape of the prostomium with a ventral cleft ( Kirkegaard, 1959) as opposed to the rounded anterior end in Myriochele ( Kirkegaard, 1959, 1983; Fauchald, 1977; Blake, 1984, 2000; Parapar, 2001, 2003b, 2006), although this genus has been considered a junior synonym of Myriochele by some authors (e.g. Blake & Dean, 1973; Nilsen & Holthe, 1985; Hartmann- Schröder, 1996; Cantone & Di Pietro, 1998; Sene- Silva, 2002). After our analyses ( Fig. 1C, D View Figure 1 ), we consider this genus as a valid taxon based on the shape of the head. We consider that the results obtained by Sene-Silva (2002) could be explained because no direct observation of specimens was carried out and the character scoring was performed from the literature (see Parapar, 2006, for further comments). Moreover, Sene-Silva’s results were incorrectly interpreted because some of the terminal taxa in his study, such as G. pygidialis and G. fragilis , were considered as members of Myriochele instead of Galathowenia ( Blake, 2000; Parapar, 2003a).
As a consequence of our results, with Myrioglobula paraphyletic and its species recovered scattered within Myriochele ( Fig. 1A–D View Figure 1 ), we propose the former as a junior synonym of the latter. Some years ago, Parapar (2003c, 2006) proposed the use of the Myriochele - Myrioglobula group after finding that the number of anterior uniramous segments showed high interspecific variability and that some species were characterized by having an alternating uniramous/ biramous pattern. Members of this clade share the presence of acicular chaetae with an elongated tapering distal end and a smooth surface, unique amongst polychaetes, and the shape of the head, similar in width to the segments and with a rounded anterior margin. Some authors have suggested Myrioglobula as a basal oweniid because of the presence of one uniramous anterior segment, a feature shared with sabellids and terebellids ( Meyer & Bartolomaeus, 1996), but this hypothesis is rejected herein and the presence of one uniramous segment is considered as a convergence in some species.
The shape of the uncini and arrangement of their teeth is a character frequently used in oweniid generic and species diagnoses. According to our phylogenetic hypothesis, this feature is informative and unambiguously characterizes Myriowenia , with teeth arranged vertically and Owenia , with side by side teeth. The teeth arranged obliquely was also one of the synapomorphies of the Galathowenia - Myriochele s.s. clade in some of the analyses ( Fig. 1D View Figure 1 ), but this was not the case either in the nonweighted data set analyses or when the concavity values were low ( Fig. 1A–C View Figure 1 ). Most Galathowenia show teeth similar in size, displayed in an oblique arrangement, but there is one exception, G. pygidialis , not included in our analyses, with teeth in a side by side position (Hartman, 1960; Blake, 2000). Members of Myriochele s.s. bear similar-sized teeth in a vertical arrangement, with the exceptions of G. australis (transferred to this clade in the present paper) and Myrioc. heruensis , which have them in an oblique position. Similarly, all the described species within the genus Myrioglobula show all the teeth in this vertical arrangement, with only the exception of Myriog. antarctica , the type species of the genus.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.