
Pseudoscalibregma ermindae, Mendes & Paiva & Rizzo, 2024
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5399.1.2 |
|
publication LSID |
lsid:zoobank.org:pub:D5584152-81FD-40BD-8A25-F95EC106CCE8 |
|
DOI |
https://doi.org/10.5281/zenodo.10513971 |
|
persistent identifier |
https://treatment.plazi.org/id/8CD6BEB7-1EB2-4D7A-9EBD-F91C3DAC30E1 |
|
taxon LSID |
lsid:zoobank.org:act:8CD6BEB7-1EB2-4D7A-9EBD-F91C3DAC30E1 |
|
treatment provided by |
Plazi |
|
scientific name |
Pseudoscalibregma ermindae |
| status |
sp. nov. |
Pseudoscalibregma ermindae sp. nov.
Figures 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3
https://zoobank.org/NomenclaturalActs/ 8CD6BEB7-1EB2-4D7A-9EBD-F91C3DAC30E1
Type material. UERJ 9205 (Holotype): SANSED4, H10 R1 , 24°13’22.4400”S, 041°31’35.4000”W, 1,912 m, coll. 25 Jul 2019 GoogleMaps ; UERJ 9206 (Paratype): SANSED1, A9 R2 , 27°19’19.9200”S, 046°31’20.6400”W, coll. 15 Jun 2019, 1,305 m GoogleMaps .
Diagnosis. Body arenicoliform, expanded over chaetigers 5–11. Body surface covered by continuous rows of smooth annuli. Rounded prostomium, with two short and rounded horns, pointing laterally. Chaetigers 1–2 biannulated, from then chaetigers triannulated throughout. Dorsal and ventral cirri present from chaetiger 15. Spines present from chaetiger 1–2, 4–5 per fascicle, with pointed tips. Lyrate chaetae present on chaetiger 3, 4–5 per fascicle, with unequal tynes.
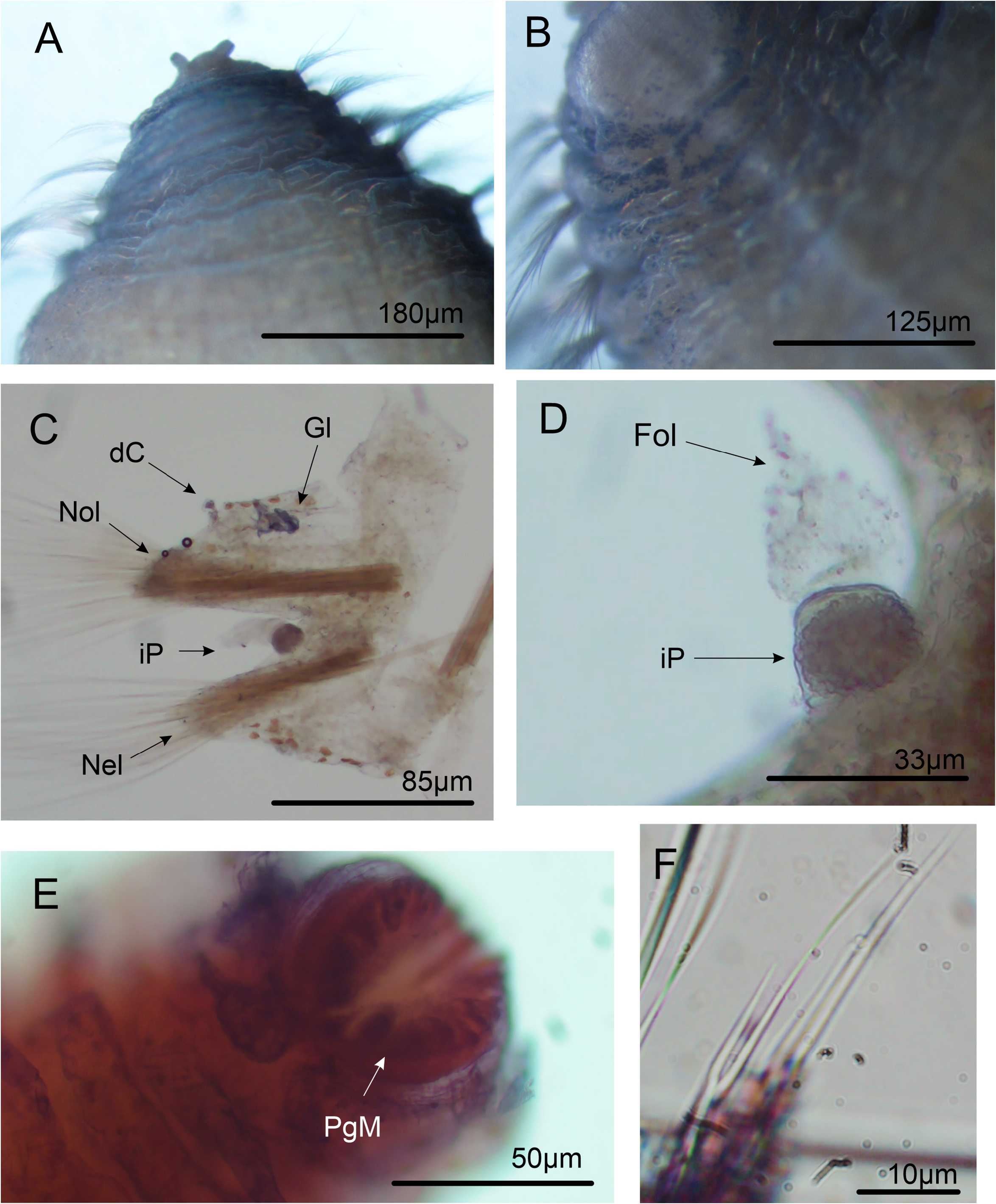
Description. Holotype complete, 20 mm long, 4 mm wide across expanded anterior region and 1.8 mm wide across narrow posterior region, with 32 chaetigerous segments. Large sized species, Paratype measuring 30 mm long to 3 mm wide across expanded anterior region and 1.5 mm wide across narrow posterior region for 33 chaetigerous segments. Body arenicoliform, expanded over chaetigers 5–11. Pale tan in alcohol. Body surface covered by continuous rows of secondary annuli; both anterior and posterior regions without conspicuous small pads on each secondary annulus; each annulus may present small individual dark blue glands within, consisting of entangled cellular tubules, located marginally along each row.
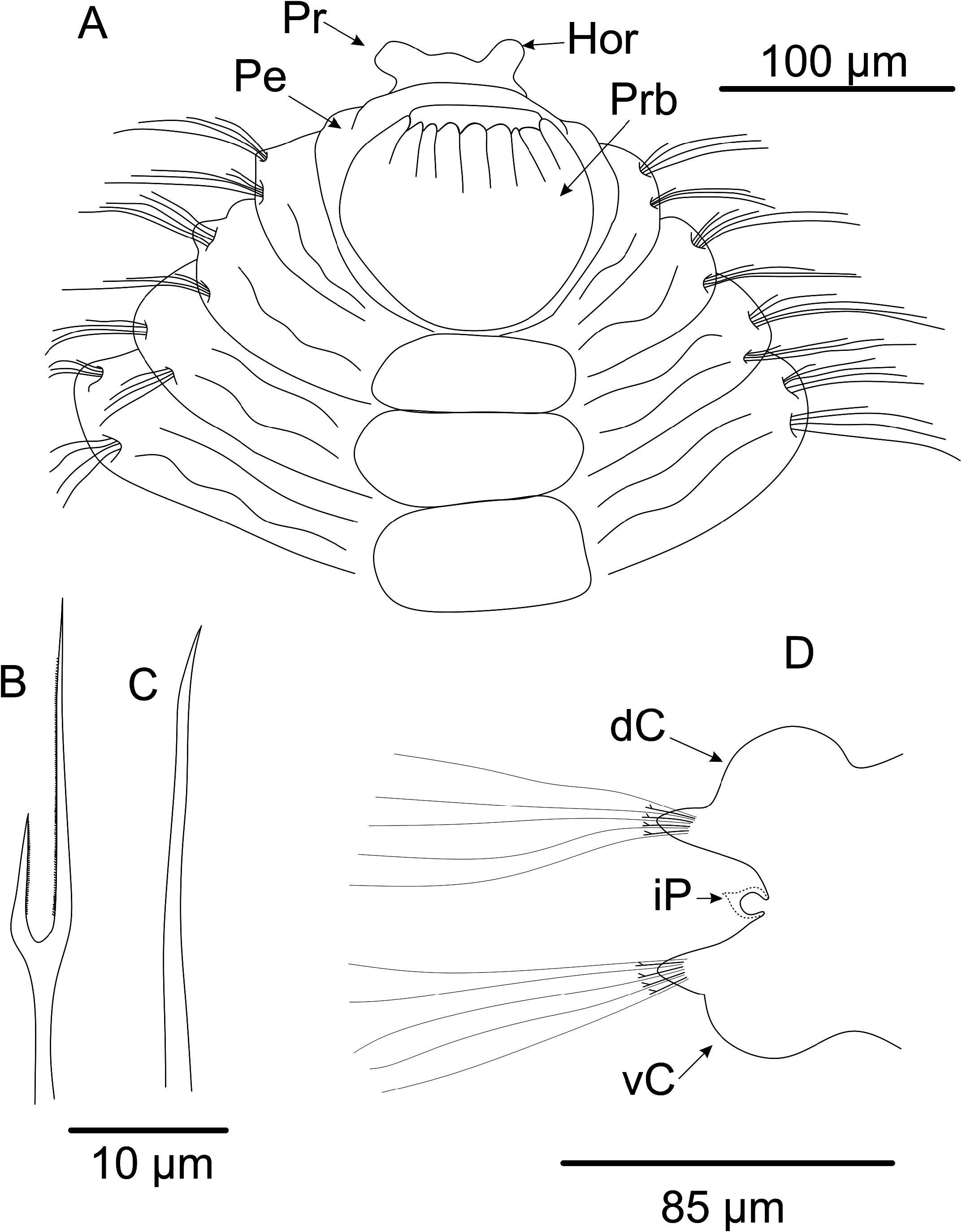
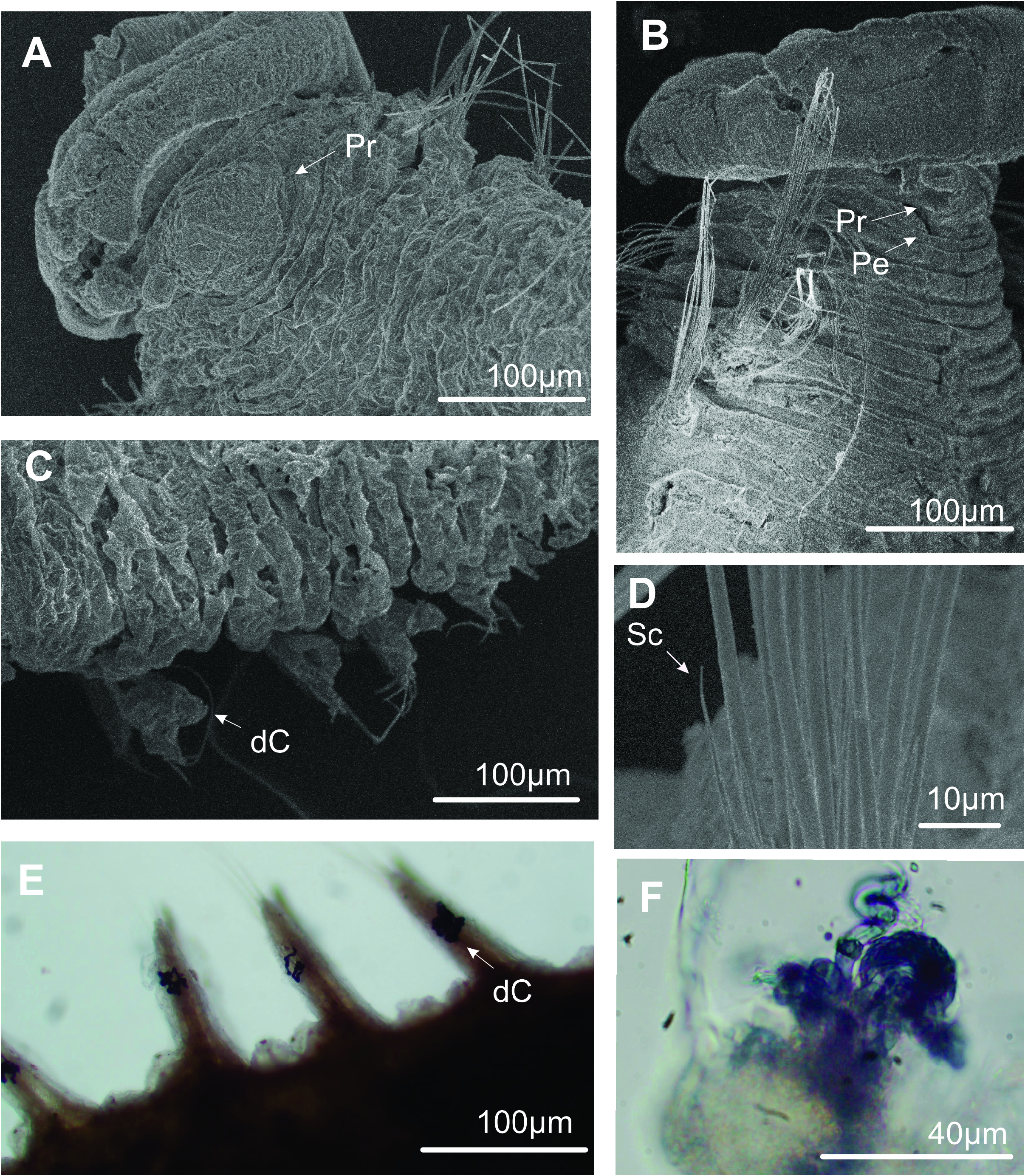
Rounded prostomium, with two short and rounded horns, both projected laterally ( Figs 1A View FIGURE 1 , 2A–B View FIGURE 2 , 3A View FIGURE 3 ). Eyes absent. Nuchal organs not observed. Peristomium achaetous, uniannulated dorsally and ventrally ( Fig. 1A View FIGURE 1 ). Proboscis smooth. Ventral groove present from chaetiger 2, with uni or biannulated rectangular pads, paired to each chaetiger ( Figs 1A View FIGURE 1 , 2B View FIGURE 2 ). Posterior pads squared to rectangular, poorly marked and smaller.
Chaetigers 1–2 biannulated, triannulated from chaetiger 3-onwards ( Figs 1A View FIGURE 1 , 2A–B View FIGURE 2 ). Dorsal and ventral cirri present from chaetiger 15; cirri blunt ( Figs 1D View FIGURE 1 , 2C View FIGURE 2 , 3C, E View FIGURE 3 ). Densely entangled dark blue to black tubular glands at midlength of cirri in all cirriferous chaetigers ( Figs 2C View FIGURE 2 , 3E–F View FIGURE 3 ). Interramal papillae present from chaetiger 15, as rounded projection with granular content within, folded by inconspicuous conical shaft ( Fig. 2C–D View FIGURE 2 ). Parapodial lobes inconspicuous on anterior chaetigers; beginning with broad base with pointed tips, forming ellipsoid-like structure, from midbody to posterior chaetigers ( Figs 1D View FIGURE 1 , 2C View FIGURE 2 ).
Short spinous chaetae present from chaetiger 1–2, 4–5 per fascicle, with pointed tips ( Figs 1C View FIGURE 1 , 3D View FIGURE 3 ). Lyrate chaetae from chaetiger 3 ( Figs 1B View FIGURE 1 , 2F View FIGURE 2 ), numbering 4–5 in each fascicle, with fine bristles and unequal tynes (tynes’ length ratio: 2.29). Capillaries in 3 rows on anterior body and 2 rows on posterior body. Pygidium structure obscure on analyzed specimens, but with a short and fusiform terminal margin, tip crenulated, lacking anal cirri ( Fig. 2E View FIGURE 2 ).
Remarks. This species is morphologically similar to P. bransfieldium ( Hartman, 1967) , P. papilia Sch ̧ller, 2008 and P. glandipodium Blake, 2023 . Pseudoscalibregma glandipodium was described from Australia, whereas P. papilia and P. bransfieldium are known to occur in South Atlantic and Antarctic waters ( Blake 1981; Scḩller 2008; Blake 2023). Pseudoscalibregma ermindae sp. nov. and P. papilia share more morphological similarities, such as a prostomium with short, rounded horns, the presence of spinous chaetae on chaetiger 1 and lyrate chaetae with unequal tynes. However, members of P. ermindae sp. nov. differ from those of P. papilia by the presence of lyrate chaetae from chaetiger 3, inflated dorsal and ventral cirri and conspicuous interramal papillae from mid to posterior chaetigers; whereas specimens of P. papilia present lyrate chaetae from chaetiger 2, foliaceous dorsal and ventral cirri and lack interramal papillae. On the other hand, P. bransfieldium is easily separated from all its congeners, including this new species, by the presence of a prominent nuchal crest on prostomium. Finally, members of P. ermindae sp. nov. can be morphologically distinguished from individuals of P. glandipodium by the presence of an interramal papillae from midbody and posterior chaetigers, and spinous chaetae on chaetigers 1–2, replaced by lyrate chaetae from chaetiger 3, whereas in specimens of P. glandipodium interramal papillae and spinous chaetae are both absent.
Etymology. The specific epithet “ ermindae ” was chosen to honour the Brazilian polychaete researcher and marine ecologist Dra. Erminda da Conceiç„o Guerreiro Couto at “Santa Cruz State University” (UESC) for her important contributions to Brazilian marine science and to celebrate our friendship.
Ecology. Apart from P. papilia and P. glandipodium , abyssal records ( Blake 2023), P. glandipodium , P. papilia , P. bransfieldium also have reports from non-abyssal depth intervals, but still deeper than shallow subtidal ranges. These records varied from 400–3,000 meters deep. The type material representing P. ermindae sp. nov. was sampled from a bathymetric range varying from 1,305 to 1,928 m deep, in muddy sediments. Hence, they also lie within the bathymetric interval of 400–3,000m shared by P. glandipodium , P. papilia , and P. bransfieldium species.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |