
Ephebopus cyanognathus West & Marshall 2000
|
publication ID |
https://doi.org/ 10.5281/zenodo.183360 |
|
DOI |
https://doi.org/10.5281/zenodo.5624766 |
|
persistent identifier |
https://treatment.plazi.org/id/03B24354-FF9E-FF94-FF44-F99DFA3BA049 |
|
treatment provided by |
Plazi |
|
scientific name |
Ephebopus cyanognathus West & Marshall 2000 |
| status |
|
Ephebopus cyanognathus West & Marshall 2000 View in CoL
( Figs 5–6 View FIGURES 1 – 10 , 16 View FIGURES 11 – 16 , 30 View FIGURE 30 )
Ephebopus cyanognathus West & Marshall 2000: 6 View in CoL , f. 1–2; West & Marshall 2002: 7.
Types: FRENCH GUIANA: Male holotype and female paratype, Roura District [4o43’ N, 52o19’ W], Tresor Mountains, Tresor Reserve, 17.VI.1999 (male matured 10.IX.1999), R. West; 2 males paratypes, Roura District [4o43’ N, 52o19’ W], Roura Mountains, 22.IV.1999 (matured in captivity 28–30.VIII.1999), R. West; 1 female, Kourou District, C.I.R.A.D. Field Station [5o09’ N, 52o38’ W], 17.IV.1999, J. Huff; 1 female, Roura District [4o43’ N, 52o19’ W], Tresor Mountains, Tresor Reserve, 17.VI.1999, R. West. All types in AMNH.
Diagnosis: Similar to E. uatuman but this species differs from all congeners by the coloration of females, the metallic blue chelicerae ( Fig. 16 View FIGURES 11 – 16 ), and the different shape of the genitalia of both sexes. Males differ from those its congeners by the palpal bulb being globular with a long slender gradually tapering embolus slightly bent outward apically ( Fig. 5 View FIGURES 1 – 10 ), similar to that in E. uatuman but straighter, thin and tapering. Spermathecae differ from those of its congeners by having two short truncate columnar lobes, separated, being widest at base and constricting medially then widening apically ( Fig. 6 View FIGURES 1 – 10 ).
Description: Holotype male, lengths: total body, 41.17; chelicerae, 7.0; carapace, 16.67; abdomen, 17.5; leg I, 72.16; leg II, 65.34; leg III, 54.5; leg IV, 68.5. Leg formula, I, IV, II, III. Color of legs dark brown; legs with narrow yellow transverse band between all femora and patellae; chelicerae with lavender pubescence; abdomen pale gold intermixed with longer gray setae; carapace and trochantera with rose and brown pubescence setae, more so on carapace. Palpal bulb globular, with long slender tapering embolus, similar to that of E. uatuman but straighter and thin, bent upward apically ( Fig. 5 View FIGURES 1 – 10 ). Paratype female, lengths: total body: 45.84; chelicerae, 6.67; carapace, 19.67; abdomen, 19.5; leg I, 65.16; leg II, 57.14; leg III, 46.20; leg IV, 58.90. Leg formula, I, IV, II, III. Color amber brown; chelicerae entirely with metallic blue pubescence; carapace lighter brown with greenish pubescence; legs and abdomen darker amber brown with narrow transverse yellow band between all femora and patellae, similar narrower transverse band of amber between all tarsi and metatarsi. Spermathecae: two shorter truncate columnar lobes, separated, widest at base, constricting medially then widening apically ( Fig. 6 View FIGURES 1 – 10 ).
Distribution. Known only from French Guiana ( Fig. 30 View FIGURE 30 ) where, in addition to the above localities, it has also been found in Maripasoula [3o43’ N, 54o04’ W] and Montsinéry District [4o54’ N, 52o30’ W], Saül [3o37’ N, 53o12’ W] and Chevaux Mountains.
Natural History. The spiders appear to be widely distributed in upland rainforest areas. They were rarely common, with the exception of a large (30+) population observed along a 10 meter long x 2 meter high road bank in Saül in September 1981 (SDM, pers. obs.). Burrows were constructed in a fully shaded laterite clay embankment, facing northeast, at about 210 meters elevation in upland rainforest. The type specimens from the Tresor Reserve were found in burrows constructed in fully shaded upland rainforest on steep, north-facing slopes in year long damp clay soil covered in leaf litter at elevations of 200–300 meters. Antepenultimate, penultimate and female E. cyanognathus construct a simple flare-mouthed tubular burrow, similar to that of E. uatuman ( Fig. 15 View FIGURES 11 – 16 ). Adult female burrows were found to terminate in an enlarged chamber about 30 cm deep (RCW and SDM, pers. obs.). We observed an adult female E. cyanognathus in her burrow with early instars in April of 2001. This specimen and her young were not collected. We have also observed an individual in an upland rainforest retreat constructed of a dead leaf in a plant in the Chevaux Mountains in March of 1999. The retreat had no obvious entrance and was about 20 cm above the ground.
Ephebopus rufescens West & Marshall 2000 View in CoL ( Figs 7–8 View FIGURES 1 – 10 , 17–18 View FIGURES 17 – 18 , 30 View FIGURE 30 )
Ephebopus rufescens West & Marshall 2000: 8 View in CoL , f. 3–4.
Types: FRENCH GUIANA: Male holotype and female paratype, Roura District, Kaw Mountains, vicinity of Camp Cäiman [4o37’ N, 51o55’ W], 17.IV.1999 (male matured 14.IX.1999), R. West & J. Huff; paratypes: 1 male, Guiana, Montsinéry District [4o54’ N, 52o30’ W], Chevaux Mountains, 17.IV.1999 (matured 18.VIII.1999), R. West & J. Huff; 1 male, Montsinéry District, Emerald Jungle Village [4o49’ N, 52o21’ W], 15.IV.1999 (matured 21.IX.1999), R. West & J. Huff; 1 female, Roura District [4o43’ N, 52o19’ W], Roura Mountains, 19.IV.1999, R. West & J. Huff; 1 female, Roura District, Kaw Mountains, vicinity of Camp Cäiman, 18.VI.1999, R. West. All types in AMNH.
Diagnosis: Females can be distinguished from those of their congeners by distinct dark reddish brown coloration and lighter narrow parallel stripes dorsally on the patellae and femora and by the lack of yellow banding on the patellal–femoral joints of all legs ( Figs 17–18 View FIGURES 17 – 18 ) and shape of the genitalia of both sexes. The male palpal bulb differs from that of its congeners by being large and globular with a shorter tapering embolus that is slightly bent apically ( Fig. 7 View FIGURES 1 – 10 ). Spermathecae are similar to those of E. uatuman but differ by being two stout separated columnar lobes ( Fig. 8 View FIGURES 1 – 10 ) that are not as wide as at base and narrowing apically as in E. uatuman .
Description: Holotype male, lengths: total body, 35.75; chelicerae, 6.42; carapace, 13.0; abdomen, 16.33; leg I, 57.99; leg II, 53.32; leg III, 45.32; leg IV, 57.17. Leg formula, I, IV, II, III. Color of legs dark brown; chelicerae, carapace, trochantera and base of all femora with longer short golden setae. Abdomen uniformly dark brown with longer reddish setae dorsally and laterally. All dorsal leg joints with narrow transverse band of short golden setae. Palpal bulb large, globular, short slender embolus tapering apically ( Fig. 7 View FIGURES 1 – 10 ). Paratype female, lengths: total body: 46.17; chelicerae, 7.33, carapace, 16.17; abdomen, 22.67; leg I, 59.16; leg II, 52.67; leg III, 44.83; leg IV, 55.0. Leg formula I, IV, II, III. Color of legs, abdomen and carapace dark reddish brown with longer lighter setae, more so on chelicerae and carapace; leg femora darker brown, dorsum of all patellae, tibiae and metatarsi with pale narrow parallel or single stripes. Spermathecae: two stout columnar lobes, separated, widest at base, similar to that in E. uatuman but not as wide at base and narrowing apically ( Fig. 8 View FIGURES 1 – 10 ).
Distribution: The type localities and French Guiana: Maripasoula [3o43’ N, 54o04’ W], Saül [3o37’ N, 53o12’ W]. Brazil: Amazonas, 30km W. of Manaus, Jacaré Creek ( Fig. 30 View FIGURE 30 ).
Natural History: The spiders were widespread and, apparently, the most ecologically versatile in their retreat placement of all Ephebopus species ( Figs 17–18 View FIGURES 17 – 18 ). Spiders were found in retreats in hollow logs and stems, both on and above the ground, in holes and hollows of standing trees, in arboreal termitaria and in mosses on the sides of shaded rock faces in rainforest. One retreat was found 3 meters above ground in a decaying palm tree. Wherever found, the spider built its retreat in association with wood or mosses. This was in or on dead or fallen trees, roots or moss covered trees, logs or rocks. A second trait of E. rufescens retreats that makes them distinct from those of other congeners is that when burrowing in the ground, the spider (particularly adult females) built an extended tubular silk retreat. This extended tubular silk retreat was generally one-quarter to one-third of the length of the burrow; it may be oriented vertically or horizontally. The retreat was usually camouflaged with soil and vegetative debris, incorporated into the silk retreat, and was attached to a woody or moss substratum. The retreat entrance had a slightly flared collar constructed of silk.
Ephebopus foliatus sp. nov.
( Figs 9–11, 14 View FIGURES 1 – 10 View FIGURES 11 – 16 , 30 View FIGURE 30 . Tables 1 View TABLE 1 , 2 View TABLE 2 )
Types: GUYANA: Male holotype and 5 paratype females, Upper Takutu–Upper Essequibo Region, 4.42 km S. Gunn’s Strip, bank of Essequibo River, 240 metres, (1°38’45.7” N, 58°38’14.6” W), 6–15.VII.1999, J.A. Coddington, G. Hormiga, J. Miller, I. Agnarsson & M. Kuntner, deposited in NMNH.
Etymology: From the Latin folium, a leaf, and refers to the subtle four-point oak leaf pattern on the dorsum of the abdomen.
Diagnosis: This species differs from all other Ephebopus by having a leaf-like pattern on the abdominal dorsum, when viewed from the side ( Fig. 14 View FIGURES 11 – 16 ), and by the shape of the genitalia of both sexes. The male palpal bulb is similar to E. cyanognathus but differs from its male congeners by having a long, thinner, tapering embolus, slightly bent apically ( Fig. 9 View FIGURES 1 – 10 ). The spermathecae differs from those of all other female congeners by having two tall columnar lobes, not widely separated, widest at base, gradually narrowing then rounding apically, sides with raised folds ( Fig. 10 View FIGURES 1 – 10 ).
Description: Holotype male, lengths: total body, 29.56; carapace, 11.36 long x 10.72 wide; abdomen, 12.0 long x 6.72 wide; chelicerae, 6.2 long x 2.28 wide; cheliceral macroteeth: (left side, right side) 10, 9 with basal granules. Sternum, 5.60 long x 4.48 wide; glabrous sigilla opposite coxae II and III, posterior sigilla the largest, 0.14 sternum width. Labium, ovoid 1.44 long x 1.92 wide; labiosternal suture a narrow groove with distinct lateral mounds; about 149 cuspules in tight cluster. Maxillae about (left, right) 180, 159 cuspules; cuspules in tight cluster on ventral inner heel of maxillae, heel rounded, anterior lobe conical and slightly elongated. Fovea: a deep circular pit. Cephalothorax: caput not rising abruptly from thoracic region but in gradual arch. Caput length 7.52, width 5.92. Ocular area 2.28 wide x 0.96 long, tubercle elevated. Anterior eye row straight; AME round, diameter 0.56, 0.23 apart; ALE elliptical, 0.33 x 0.44, 1.47 apart. Posterior eye row slightly recurved; PME ovoid, 0.21 x 0.37, 1.19 apart; PLE ovoid, 0.28 x 0.48, 1.49 apart. Clypeus 0.48 wide. Leg span 112.0mm, taken on right side (leg IV missing. Leg formula I, IV, II, III ( Table 1 View TABLE 1 ). Femur III not noticeably swollen when viewed from above. All tarsal scopulae entire, without setal division. Metatarsal scopulae: I, 0.77; II, 0.85; III, 0.67; IV, 0.28. Type V urticating hairs in dense pad on apical prolateral face of palpal femur. No plumose setae on palp or leg segments. Spination: leg I, metatarsus 1v (1ma), tibia 4v (3 megaspines, one along entire inner lower process length, two on apical upper process, one on inner and one on outer, 1 ra); leg II, metatarsus 1v (1ma), tibia 2v (1pa, 1ra); leg III, metatarsus 3v (1pa, 1ma, 1ra), tibia, 2v (1pa, 1ra);;leg IV, metatarsus 2v (1pa, 1ra); palp, aspinose. Spur on tibia I bipartite; lower process 1.20 long, with single basal megaspine along entire inner face; upper process 1.80 long, with preapical megaspine on opposing inner and outer curved face. Metatarsus I closes on outer upper process. Palpal bulb similar to E. cyanoganthus but globular, with long thinner tapering embolus, slightly bent apically ( Fig. 9 View FIGURES 1 – 10 ). Color (in alcohol): carapace golden brown woolly pubescence; abdomen golden brown with longer reddish setae; chelicerae golden brown; legs brown with narrow transverse yellow band between all femora and patellae. Paratype female, lengths: total body: 38.08; carapace, 14.88 long x 12.8 wide; abdomen, 16.32 long x 12.0 wide; chelicerae, 6.88 long x 3.36 wide; cheliceral teeth: (left side, right side) 10,13. Sternum: length 6.24, width 5.4; glabrous sigilla opposite coxae II and III, posterior sigilla the largest, 0.15 of sternum width. Labium: 2.46 long, 2.04 wide; labiosternal suture with distinct lateral mounds; ca. 198 cuspules in tight cluster; maxillae with ca. 184–207 cuspules. Fovea straight, deep. Cephalothorax: caput not rising abruptly from thoracic region but in gradual arch. Caput length 9.12, width 8.0. Ocular area 2.64 wide x 1.20 long, tubercle relatively low. Anterior eye row straight; AME round, diameter 0.67, apart 0.30; ALE elliptical, 0.26 x 0.58, apart 1.84. Posterior eye row slightly recurved; PME ovoid, 0.21 x 0.37, apart 1.51; PLE elliptical, 0.23 x 0.44, apart 1.98. Clypeus 0.47 wide. Leg span 105.0 mm. Leg formula I, IV, II, III ( Table 2 View TABLE 2 ). All tarsal scopulae entire, without setal division. Metatarsal scopulae: I – II, entire; III, 0.73; IV, 0.44. Type V urticating hairs in dense pad on apical prolateral face of palpal femur. No plumose setae on palp or leg segments. Spination: Leg I, tibia 2v (1pa, 1ra); II, metatarsus 1v (ma), tibia 1v (pa); III, metatarsus 3v (1pa, 1ma, 1ra), tibia 2v (1pa, 1ra); IV, metatarsus 2v (1pa, 1ra), tibia 2v (1pa, 1ra); palp 4v (2pa, 2ra). Spermathecae, two tall columnar lobes, not widely separated, widest at base, abruptly narrowing then rounded apically, with raised folds ( Fig. 10 View FIGURES 1 – 10 ). Color: (in alcohol): chelicerae, carapace and abdomen rufus brown with scattered longer reddish brown setae on the abdomen; legs darker rufus brown with narrow transverse yellow bands between all leg joints, widest between femora and patellae.
Variation: Males: only the holotype. Females: 5 (including the paratype), length 29.28–36.16; carapace length 11.68–14.88, width 10.72–12.8; carapace width/length 0.86–0.92; chelicerae length 5.6–6.88, width 2.88–3.36; abdomen length 11.68–17.12, width 8.48–13.12; leg span 95.0–105.0 mm. Cheliceral teeth 9,10– 11,13. Metatarsal scopulae: I–II, entire; III, 0.70–0.77 (0.74); IV, 0.34–0.44 (0.39). Type V urticating hairs in dense pad on apical prolateral face of palpal femur. No plumose setae on palp or leg segments. Spermathecae, as in paratype.
Distribution: Known only from type locality in Guyana ( Fig. 30 View FIGURE 30 ).
Natural History: All specimens were collected at night from 1–4 m above ground on the sides of trees in riparian rainforest ( Fig. 14 View FIGURES 11 – 16 ). None were found on the ground or observed in silken retreats. We lack the field observations on the natural history of E. foliatus , however, collecting records indicate this species is arboreal (M. Kuntner, per. comm., USNM, 2002).
Characters: In characters marked with an asterisk (*; viz., 6, 10, 12, 13, 18, 20, 31, 39, 42), fit and steps are not given as the character is not significant to the analysis but was kept here to indicate an autapomorphy for a terminal taxon. (0) Eye tubercle in males and females: (0) low ( Fig. 23 View FIGURES 23 – 26 ), (1) high ( Fig. 24 View FIGURES 23 – 26 ); fit = 66.6. Steps/ Extra steps = 4/3. The eye tubercle can be absent in some mygalomorphs (e.g., Atypidae, Rastelloidina, Raven 1985 ; Goloboff 1993b, 1995) or well-defined in most mygalomorph taxa. Intermediate states have been proposed for some cyrtaucheniids, hexathelids, ctenizids ( Raven 1985:125, 70, 141; Bond & Opell 2002), and diplurids ( Ischnothele Ausserer ) ( Bond & Opell 2002). In theraphosids, two states have been proposed: absent, for some Hemirrhagus Simon 1903 , ( Spelopelma Gertsch ; Raven 1985) and distinct, for remaining species. An intermediate state was herein identified: Heteroscodra ( Fig. 23 View FIGURES 23 – 26 ) shows a somewhat flattened eye tubercle, with anterior lateral eyes being in almost same plane as anterior median eyes (state 0). Spiders with an arched eye tubercle ( Fig. 24 View FIGURES 23 – 26 ), with the anterior median eye positioned clearly in superior plane in relation to the anterior lateral eye were considered as having state 1.
(1) Anterior row of eyes in males and females, measured from anterior margins: (0) procurved, (1) straight; fit = 85.7. Steps/ Extra steps = 2/1.
(2) Clypeus in males and females: (0) absent, (1) narrow ( Fig. 23 View FIGURES 23 – 26 ) (2) wide ( Fig. 24 View FIGURES 23 – 26 ); fit = 60.0. Steps/ Extra steps = 6/4. The clypeus was considered absent when there is no space between the eye tubercle and the anterior carapace edge which can slightly project forward beyond the anterior carapace edge. A wide clypeus is found in some theraphosids, e.g. Harpactirinae , Ornithoctoninae and some theraphosines. In this case, the distance between the anterior edge of the eye tubercle and the anterior edge of carapace can be almost the same as the length of the eye tubercle ( Fig. 24 View FIGURES 23 – 26 ). The narrow state of the clypeus occurs in intermediate cases ( Fig. 23 View FIGURES 23 – 26 ).
(3) Fovea in males and females, curvature: (0) straight, (1) recurved, (2) procurved; fit = 85.7. Steps/ Extra steps = 3/1.
(4) Fovea in males and females, closure: (0) slit-like (closed), (1) pit-like (open); fit = 100. Steps/ Extra steps = 1/0. In Mygalomorphae, a pit-like (open) fovea seems to be plesiomorphic, since Mesothelae shows this state as well as representatives of Atypidae , Antrodiaetidae and Dipluridae ( Raven 1985) . For theraphosids, Gallon (2003) suggested the presence of pit-like fovea to be a synapomorphy of the clade Xenodendrophila Gallon 2003 (now Encyocratella Strand ) ( Stromatopelma + Heteroscodra ). This proposal is followed here.
(5) Fovea in males and females, depth: (0) shallow; (1) deep; fit = 66.6. Steps/ Extra steps = 4/3. This character is introduced here and refers to the depth of the fovea. Plaster was applied to the specimen's fovea and the resulting mould was analyzed. Those with deep fovea (state 1) presented a mould with trapezoid shape. Specimens with shallow fovea (state 0) showed a mould with a short line.
(6) Labial cuspules in males and females, number*: (0) 30–300, (1) 0–20, (2) 350–450 Within the Theraphosidae examined here, only Euathlus has a low number of labial cuspules. The most common state is between 30-300 labial cuspules and the only taxon with more than this number is Phlogiellus . The state 30– 300 was coded as 0 because it is present in the outgroups and thus should be the plesiomorphic state.
(7) Sigilla, posterior pair in males and females, position: (0) marginal, less than 1.5 times its own diameter from margin, (1) located closer to center of sternum separated from margin by twice its own diameter; fit = 75. Steps/ Extra steps = 3/2. In Eumenophorinae, the posterior sternal sigilla are medially placed ( Pocock 1897; Gallon 2003). We consider Phlogiellus and Haplopelma also have this state.
(8) Tarsus IV in males, cracked: (0) cracked, (1) integral; fit = 85.7. Steps/ Extra steps = 2/1. A cracked tarsus is found in some mygalomorph taxa, e.g., some diplurids, nemesiids, barychelids and theraphosids ( Raven 1985). In specimens examined, this condition was found in males and females of Melloina and males of Holothele and Phlogiellus .
(9) Tarsal scopulae in males and females: (0) no true scopula; (1) scopula of sparse hairs; (2) dense scopula that does not extend much laterally; (3) scopula very extensive laterally, giving the tarsi and metatarsi I and II a spatulate appearance; fit = 85.7. Steps/ Extra steps = 4/1. Melloina has no true scopula (state 0) ( Raven 1999). In Holothele and other Ischnocolinae, the scopula is very sparse (state 1), and frequently divided by rows of setae. Most theraphosids have well-developed scopulae (state 2), that, however, is not so developed as those having state 3, where their lateral development give the tarsi and the metatarsi I and II a spatulate appearance. This last state was used by Simon (1892) to characterize the Avicularieae and that was followed by most authors thereafter.
(10) Scopulae on metatarsus IV in males and females*: (0) divided by hairs or spines; (1) not divided. Gallon (2003) considered the undivided scopula on metatarsus IV to be apomorphic for Eumenophorinae. The only taxon with the derived state in the analysis is Citharischius Pocock 1901 .
(11) Leg spines in males and females: (0) present on apex and other faces of tibiae and metatarsi, (1) present only on ventral apices of tibiae and metatarsi, (2) absent; fit = 75. Steps/ Extra steps = 4/2. The theraphosid sister-groups, e. g., Melloina and barychelids, have spines distributed largely on ventral, prolateral, retrolateral and sometimes dorsal faces of the legs. This suggests that the condition is plesiomorphic for Theraphosidae . Among the studied taxa, a lower number of leg spines is seen in Tapinauchenius , Psalmopoeus , Ephebopus , Citharischius and Phlogiellus where there are only two small spines on the ventral apical edge of the tibiae and metatarsi. In Avicularia , Iridopelma , Pachistopelma , Stromatopelma , Heteroscodra and Poecilotheria , spines are totally absent.
(12) Palpal femora in males and females, scopula on retrolateral face*: (0) absent, (1) present. Raven (1985) included Stromatopelma in the Eumenophorinae because they share the presence of a brush of hair on the retrolateral face of the palpal femora. Gallon (2003) stated that only Stromatopelma females present this scopulae and proposed its presence to be homoplastic with respect to Eumenophorinae. We failed to find this structure in males and females of Stromatopelma examined, and so it was here coded as absent. The only genus in the analysis clearly having the scopulae was Citharischius .
(13) Femora IV in males and females, scopulae on retrolateral face*: (0) absent, (1) present.The scopulae on the retrolateral face of femora IV is an apomorphy of Lasiodora C. L. Koch 1850 and other theraphosine genera not included in the analysis ( Pérez-Miles et al. 1996).
(14) Chelicerae in males and females, scopulae on retrolateral face: (0) absent, (1) present; fit = 85.7. Steps/ Extra steps = 2/1.
(15) Maxillae, spiniform setae on lower prolateral face: (0) absent, (1) present; fit = 100. Steps/ Extra steps = 1/0. Spiniform setae on the lower prolateral face of maxillae are found in Ornithoctoninae and Thrigmopoeinae ( Raven 1985). We here consider that spines in the same position in Poecilotheria are putative primary homologs.
(16) Stridulatory bristles in males and females form maxillae lyra: (0) absent, (1) present; fit = 75. Steps/ Extra steps = 3/2. The derived condition (1) of this character was used to characterize the Selenocosmiinae (e.g., Pocock 1895). It is also found in other mygalomorph taxa apart from Theraphosidae , e.g., the diplurids Diplura C. L. Koch 1850 and Trechona C. L. Koch 1850 ( Raven 1985) .
(17) Stridulatory bristles on coxae I of males and females: (0) absent, (1) present; fit = 85.7. Steps/ Extra steps = 2/1. Stridulatory bristles on coxae were used to characterize the Eumenophorinae (e.g., Raven 1985) and some Theraphosinae genera such as Lasiodora , Grammostola Simon 1892 and Theraphosa Thorell 1870 ( Pérez-Miles et al. 1996) .
(18) Longitudinal white stripes on patellae and tibiae of males and females*: (0) absent, (1) present ( Fig. 11 View FIGURES 11 – 16 ). The conspicuous white stripes in Ephebopus murinus were considered an autapomophy.
(19) Leg rings on distal femora, tibiae and metatarsi of males and females: (0) absent, (1) white, (2), yellow ( Figs 13–14, 16 View FIGURES 11 – 16 ); fit = 85.7. Steps/ Extra steps = 3/1. Most theraphosid species have a band of short setae around the distal femora, tibiae and metatarsi of all legs, more visible on legs I and II. Some taxa, e. g. Holothele , have no detectable rings, whereas in most theraphosids, they are white, if present. Some species of Ephebopus and Avicularia have these rings but are yellow in color.
(20) Chelicerae of females, hair: (0) not iridescent, (1) iridescent ( Fig. 16 View FIGURES 11 – 16 ). State 1 is an autapomophy of E. cyanognathus .
(21) Black marking dorsally on tibiae, metatarsi and tarsi of males and females: (0) absent, (1) present; fit = 100. Steps/ Extra steps = 1/0. This character was introduced by Gallon (2003) and his proposal is followed here.
(22) Female abdominal pattern: (0) all one color or irregularly mottled, (1) with pattern; fit = 60. Steps/ Extra steps = 5/4. Gallon (2003) considered that ornithoctonines, harpactirines and "stromatopelmines" ( Heteroscodra , Stromatopelma and Xenodendrophila , now Encyocratella ) to have abdomen dorsums marked with a herring-bone pattern. We follow that here and include also Poecilotheria and E. foliatus as well as Avicularia , Pachistopelma , Iridopelma , Tapinauchenius and Psalmopoeus Pocock 1895 in which genera, immatures have a dorsal abdominal pattern.
(23) Spermathecae, number: (0) two, completely separated, (1), two, fused at base, (2) one, totally fused; fit = 100. Steps/ Extra steps = 2/0.
(24) Spermathecal lobes: (0) absent, (1) present; fit = 66.6. Steps/ Extra steps = 4/3.
(25) Cymbium: (0) without spiniform process between lobes, (1) with spiniform process between lobes; fit = 100. Steps/ Extra steps = 1/0. This character was proposed by Raven 1985 as a putative synapomorphy of Aviculariinae .
(26) Prolateral cymbium lobe shape: (0) rounded ( Fig. 26 View FIGURES 23 – 26 ), (1) subtriangular ( Fig. 25 View FIGURES 23 – 26 ); fit = 85.7. Steps/ Extra steps = 2/1.
(27) Subtegulum: (0) small, (1) large, extending down the bulb for half of the tegulum ( Raven 1985); fit = 85.7. Steps/ Extra steps = 2/1. A large subtegulum is considered a Theraphosinae synapomorphy ( Raven 1985; Pérez-Miles et al. 1996). This character state 1 is also present in Haplopelma Simon 1892 .
(28) Embolus curvature: (0) straight ( Figs 1, 3, 5, 7, 9 View FIGURES 1 – 10 ), (1) curved to the retrolateral side ( Fig. 25 View FIGURES 23 – 26 ); fit = 66.6. Steps/ Extra steps = 4/3.
(29) Embolus length: (0) 1.5-2.5 times longer than tegulum, (1) shorter than tegulum, (2) more than 3 times longer than tegulum; fit = 60. Steps/ Extra steps = 6/4.
(30) Embolus distal width: (0) thin, less than 1/5 of tegulum height ( Fig. 25 View FIGURES 23 – 26 ), (1) thick, more than 1/3 of tegulum height ( Fig. 26 View FIGURES 23 – 26 ); fit = 85.7. Steps/ Extra steps =2/1
(31) Embolus shape*: (0) not flattened, (1) slightly flattened, (2) very flattened. The states follow Bertani (2000; 2001). Haplopelma was found to have a very flattened embolus, similar to some theraphosine species, such as Theraphosa , Megaphobema Pocock 1901 and Sericopelma Ausserer 1875 .
(32) Prolateral inferior keel of bulb: (0) absent, (1) present; fit = 100. Steps/ Extra steps =1/0. Several mygalomorph taxa have strongly developed keels on the male palpal bulb, e.g., species of Pycnothelinae ( Nemesiidae ), Barychelinae ( Barychelidae ) and Theraphosinae ( Raven 1985) . In Theraphosidae , the presence of keels was considered a synapomorphy of Theraphosinae ( Raven 1985; Pérez-Miles et al. 1996). Bertani (2000) created a terminology for theraphosine male palpal bulb keels, which is followed here. However, other theraphosid taxa have strongly developed keels, such as species of Haplopelma and Poecilotheria . These keels are similar in position to some theraphosine species and thus they are being putatively considered primary homologs in the cladistic analysis.
(33) Prolateral superior keel on embolus ( Bertani 2000): (0) absent, (1) present; fit = 100. Steps/ Extra steps =1/0. (See character 32).
(34) Apical keel on embolus ( Bertani 2000): (0) absent, (1) present; fit = 85.7. Steps/ Extra steps = 2/1. (See character 32).
(35) Retrolateral keel on embolus ( Bertani 2000): (0) absent, (1) present; fit = 85.7. Steps/ Extra steps = 2/ 1. (See character 32).
(36) Male tibial spurs: (0) present, two-branched, (1) present, with apical megaspine, (2) present, with spiniform setae, (3) absent; fit = 66.6. Steps/ Extra steps = 6/3. Male tibial spurs are a very homoplasious character in Theraphosidae ( Bertani 2001) and most other mygalomorph families ( Raven 1985). However, the presence of two-branched spurs seems to be plesiomorphic in Theraphosidae since it is present in all barychelid subfamilies and in Melloina (Paratropididae) , the proposed sister-groups of Theraphosidae ( Raven 1985) .
(37) Hairs on metatarsi and tibiae I–IV of males: (0) normal, (1) long hairs laterally projected, forming a brush; fit = 85.7. Steps/ Extra steps = 2/1. Males of Tapinauchenius , Psalmopoeus , Stromatopelma and Heteroscodra all share the presence of hairs projecting laterally.
(38) Urticating hairs of males and females: (0) absent, (1) on abdomen ( Cooke et al. 1972), (2) on pedipalps ( Marshall & Uetz 1990) ( Fig. 19 View FIGURES 19 – 20 ). Fit = 85.7. Steps/ Extra steps = 3/1.
(39) Type I urticating hairs in males and females*: (0) absent, (1) present.
(40) Type II urticating hairs in males and females: (0) absent, (1) present; fit = 100. Steps/ Extra steps = 1/ 0.
(41) Type III urticating hairs in males and females: (0) absent, (1) present; fit = 100. Steps/ Extra steps = 1/0. Considered a synapomorphy of Theraphosinae ( Pérez-Miles et al. 1996).
(42) Type IV urticating hairs in males and females*: (0) absent, (1) present.
(43) Type V urticating hairs in males and females: (0) absent, (1) present ( Figs 19–20 View FIGURES 19 – 20 ); fit = 100. Steps/ Extra steps = 1/0. Type V hair ( Fig. 20 View FIGURES 19 – 20 ) is present in Ephebopus murinus ( Marshall 1990) and all known species of this genus.
(44) Burrow entrance (fossorial) of female: (0) simple with little or no silk, (1) collar of silk bound with surrounding debris ( Fig. 15 View FIGURES 11 – 16 ), (2) trumpet-shaped ( Fig. 11 View FIGURES 11 – 16 ); fit = 85.7. Steps / Extra steps = 3/1.
(45) Eggsac type: (0) mobile, (1) fixed hammock, (2) fixed flat; fit = 100. Steps / Extra steps = 2/0. Here we follow the classification of Gallon & Gabriel (2006)
(46) Habitat: (0) evergreen forest, (1) deciduous forest, (2) desert-scrub; fit = 85.7. Steps/ Extra steps = 3/ 1.
(47) Habits of female: (0) hide in surface layers of soil, (1) arboreal, (2) opportunistic ( Figs 17–18 View FIGURES 17 – 18 ), (3) fossorial ( Fig. 11 View FIGURES 11 – 16 ). Fit = 75. Steps / Extra steps = 5/2. State 0 was reported in paratropidids ( Raven 1999).
Most theraphosids make a burrow (state 3) or hide in retreats built in trees using silk (most aviculariines and Poecilotheria ) (state 1). An opportunistic burrowing behavior (state 2) is found in Tapinauchenius , Psalmopoeus and Ephebopus rufescens (RCW and SDM pers. obs.). They build a silken retreat which is not always made high in trees or other elevated structures. Furthermore, they commonly use soil and/or rocky substrata.
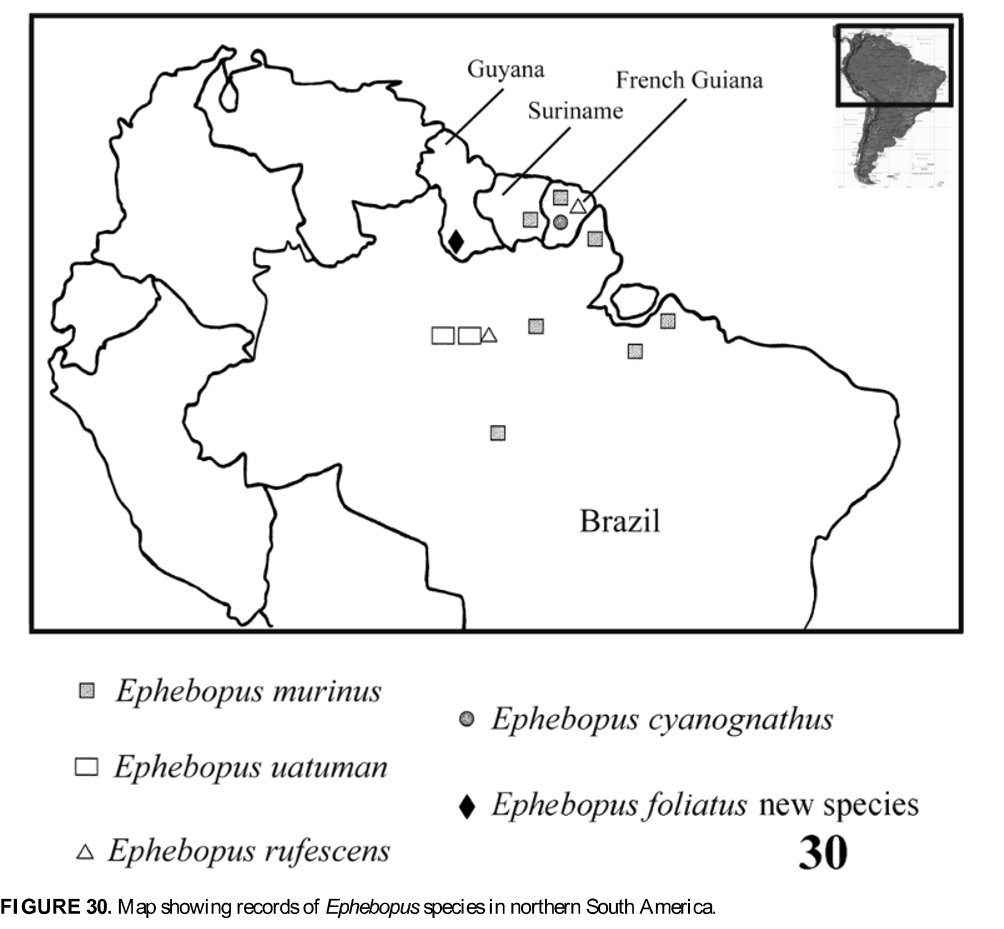
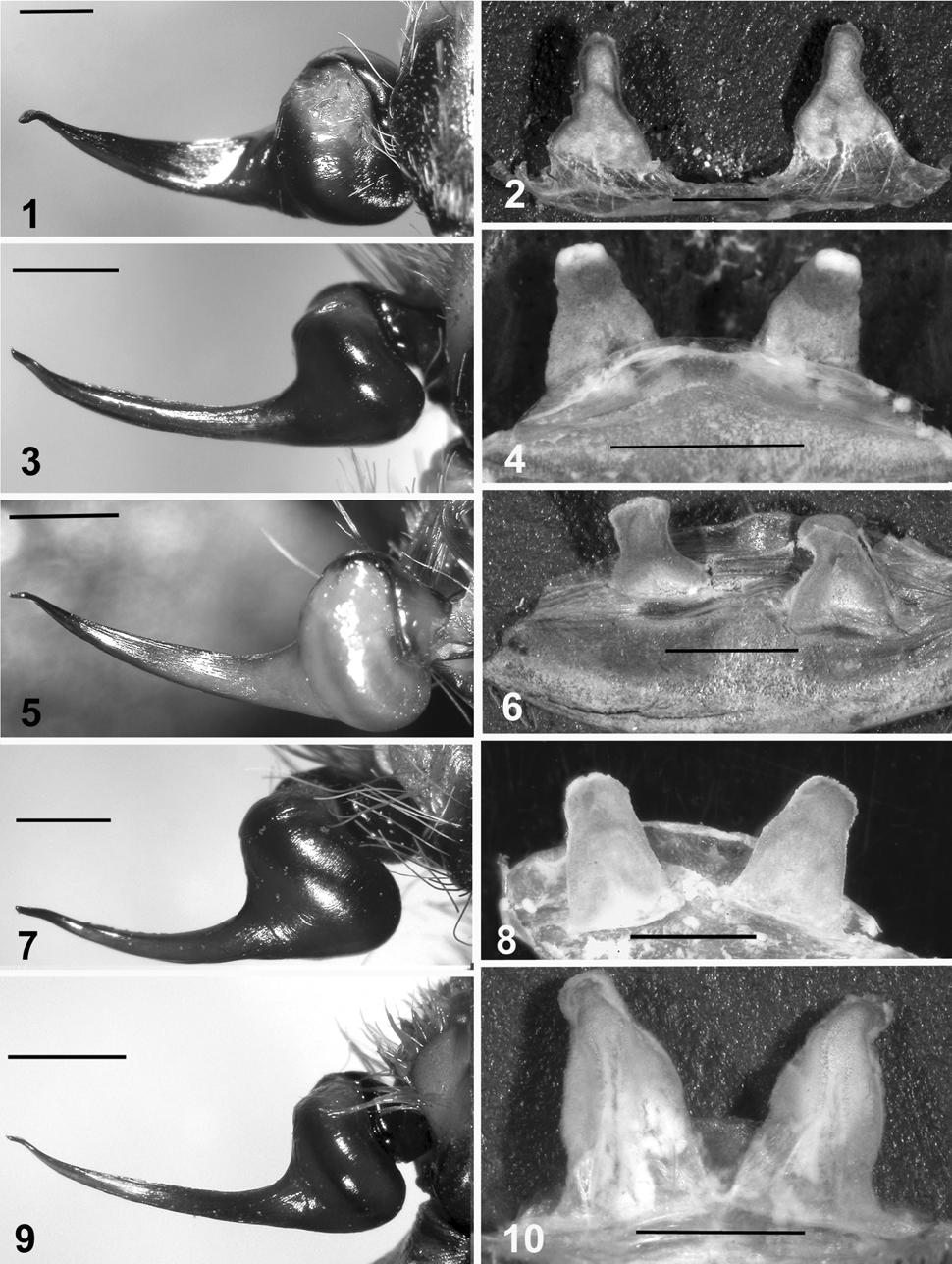


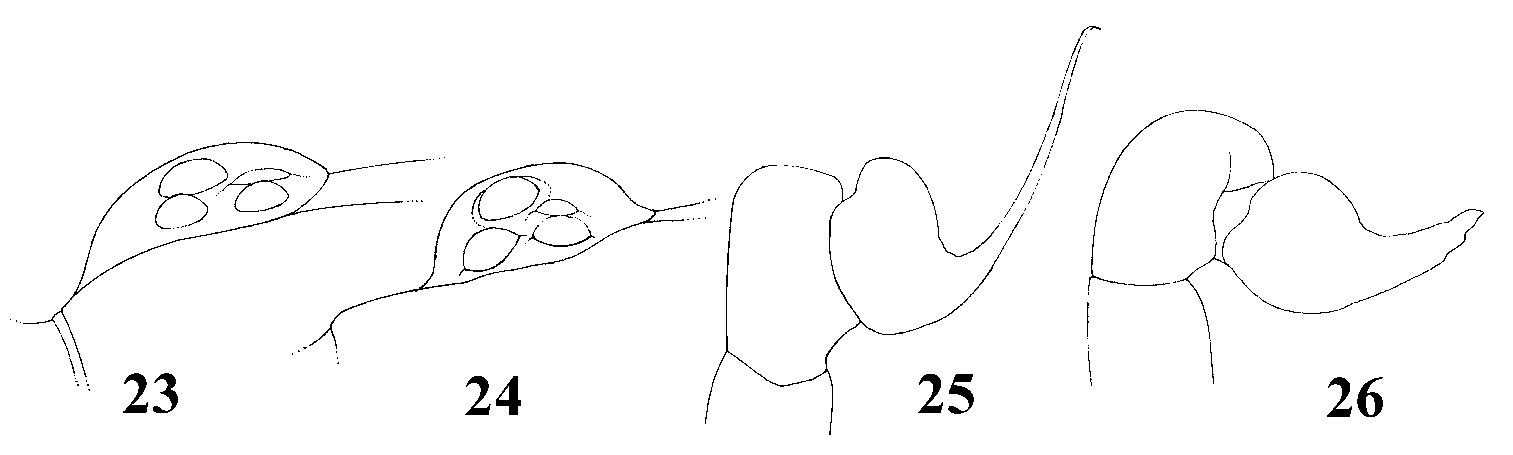

TABLE 1. Leg and palp segment lengths in mm of Ephebopus foliatus, holotype male.
| coxae | trochanter | femur | patella | tibia | metatarsus | tarsus | total | |
|---|---|---|---|---|---|---|---|---|
| palp | 4.0 | 2.4 | 7.2 | 2.72 | 6.88 | - | 2.56 | 25.76 |
| legI | 5.28 | 3.2 | 13.76 | 7.04 | 12.16 | 11.2 | 5.12 | 57.76 |
| legII | 4.32 | 3.04 | 12.0 | 6.08 | 10.08 | 10.4 | 4.96 | 50.88 |
| legIII | 3.68 | 2.4 | 9.92 | 4.8 | 8.0 | 10.56 | 4.32 | 43.68 |
| legIV | 4.0 | 2.88 | 12.64 | 5.28 | 11.2 | 14.4 | 4.32 | 54.72 |
TABLE 2. Leg and palp segment lengths in mm for paratype female of Ephebopus foliatus.
| coxae | trochanter | femur | patella | tibia | metatarsus | tarsus | total |
|---|---|---|---|---|---|---|---|
| palp 4.96 | 3.68 | 8.8 | 4.96 | 5.44 | - | 5.6 | 33.44 |
| Leg I 6.4 | 3.2 | 12.48 | 7.36 | 9.6 | 8.96 | 4.8 | 52.8 |
| Leg II 5.6 | 3.36 | 11.2 | 6.72 | 7.84 | 8.32 | 4.48 | 47.52 |
| Leg III 3.84 | 2.88 | 9.12 | 5.12 | 9.12 | 8.0 | 4.16 | 42.24 |
| Leg IV 5.28 | 3.2 | 11.52 | 5.6 | 9.44 | 11.62 | 4.16 | 50.88 |
| Cladistics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Ephebopus cyanognathus West & Marshall 2000
| West, Rick C., Marshall, Samuel D., Fukushima, Caroline Sayuri & Bertani, Rogério 2008 |
Ephebopus cyanognathus West & Marshall 2000 : 6
| West 2002: 7 |
| West 2000: 6 |
Ephebopus rufescens West & Marshall 2000 : 8
| West 2000: 8 |