
Macellicephala monroi, Neal & Brasier & Wiklund, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4455.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:70E92EDF-E11B-40A7-9573-0AC9F10E623E |
|
DOI |
https://doi.org/10.5281/zenodo.5994479 |
|
persistent identifier |
https://treatment.plazi.org/id/03BDA104-A328-694C-FF64-FD1EFBDA24B5 |
|
treatment provided by |
Plazi |
|
scientific name |
Macellicephala monroi |
| status |
sp. nov. |
Macellicephala monroi View in CoL sp. nov.
( Figure 2. C, D View FIGURE 2 and Figures 7 View FIGURE 7 , 8 View FIGURE 8 ; Table 2.)
Macellicephala View in CoL sp. in Monro (1930): 47, p. 47: Fig. 10a, b View FIGURE 10 [description].
Macellicephala mirabilis in Neal et al. (2017) View in CoL [record].
Not Macellicephala mirabilis in Monro (1936): 100 View in CoL [description].
Material examined. Holotype, NHMUK.2018.53, South Georgia (Scotia Sea, Southern Ocean ), cruise JR 144, station SG-EBS-3, collected on 05/04/2006, epibenthic sledge, epi-net, 53°35'51''S, 37°54'7.632''W, 503 m depth GoogleMaps . Paratype, historic specimen of Monro (1930), BMNH 1930 :10.8.440, South Georgia, Discovery, collected on 15/ 12/1926, 230– 250 m depth. DNA voucher, NHMUK.2018.75, Powell Basin, ( Scotia Sea , Southern Ocean ), cruise JR 144, station PB-EBS-3, collected on 18/03/2006, epibenthic sledge, epi-net, 60°59'24.72''S, 46°49' 54.516''W, 504 m depth. GoogleMaps
Additional material examined: Two specimens of Monro (1936), BMNH 1936 : 2: 8: 488, South Georgia (mouth of Stromness Harbour), Discovery station 144, 155– 178 m depth.
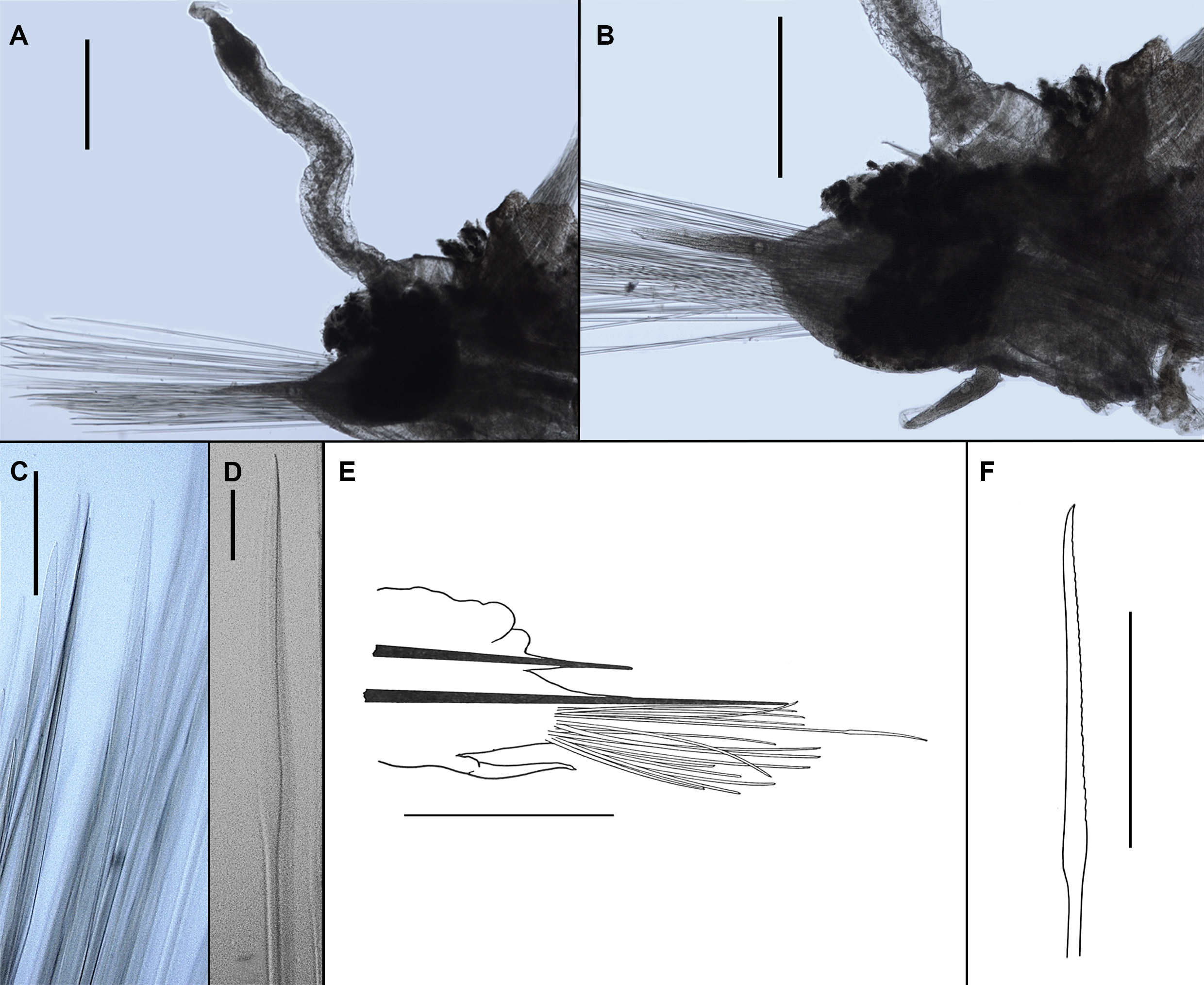
Description (based on holotype and paratype). Holotype complete and in overall good condition, with many body appendages still attached ( Figs. 2D View FIGURE 2 , 7A View FIGURE 7 ). Paratype, historic specimen of Monro (1930) ( Figs. 2C View FIGURE 2 , 7C View FIGURE 7 ), styles of median antenna, tentacular and dorsal cirri missing. Holotype 8 mm long, 2 mm wide excluding parapodia and 4 mm wide including parapodia; with 18 segments (segment 1 = tentacular segment); body dorsoventrally flattened, tapering anteriorly and posteriorly; colour in alcohol pale yellow with distinct pigmentation of purple and rusty brown stripes horizontally across dorsum observed in all specimens examined ( Fig. 7A, C View FIGURE 7 ).
Prostomium bilobed with anterior notch; prostomial lobes slightly pronounced, anteriorly tapering. Frontal filaments extending from tapering lobes short, medially inserted ( Figs. 2C, D View FIGURE 2 and 7B View FIGURE 7 ). Eyes absent. Facial tubercle indistinct, only present as enlarged inflated pad. Ceratophore of median antenna prominent, short, cylindrical, inserted medially on prostomium (near the base of anterior notch); style of median antenna robust, somewhat inflated, smooth, similar in length to palps ( Figs. 2D View FIGURE 2 , 7B View FIGURE 7 ). Palps thick, tapered, smooth, reaching to about segment 4.
Tentaculophores cylindrical, of equal size, inserted laterally to prostomium, achaetous; styles similar in form to style of median antenna ( Fig. 2D View FIGURE 2 ), with dorsal tentacular style longer than ventral style (ventral tentacular style about 2/3 length of dorsal style). Pharynx not observed. Second segment with elytrophores, biramous parapodia, chaetae and ventral cirri.
Nine pairs of prominent, bulbous elytrophores ( Fig. 7D View FIGURE 7 ), on segments 2, 4, 5, 7, 9, 11, 13, 15 and 17; all elytra missing. Dorsal ridges absent.
Parapodia biramous. Notopodia reduced to elongate acicular lobe ( Fig. 8B, E View FIGURE 8 ), tip of notoacicula not penetrating epidermis. Neuropodia large ( Fig. 8B, E View FIGURE 8 ), subtriangular tapering to elongate acicular lobe, tip of neuroacicula not penetrating epidermis. Cirrigerous segments with prominent bulbous to inflated cirrophores, inserted basally on notopodia ( Figs. 7D View FIGURE 7 , 8A, B View FIGURE 8 ); styles robust ( Figs. 7D View FIGURE 7 , 8A View FIGURE 8 ), somewhat inflated, smooth, tapering into slender tip, long (exceeding neuropodial acicular lobe), similar in form to styles of median antenna and tentacular cirri. Inconspicuous dorsal tubercles present, best observed on segments 3 and 6 as a small conical mound then becoming indistinct. Ventral cirri smooth, approaching bottle-shaped form (with slightly enlarged base than tapering), present from segment 2; inserted basally on segment 2, where very long, (longer than associated neuropodial lobe); inserted medially on subsequent segments ( Fig 7B View FIGURE 7 ), where short (not reaching distal margin of neuropodial lobe).
Notochaetae missing (only broken stumps observed). Neurochaetae numerous, translucent, upper and lower neurochaetae shorter, others very long; distal half of chaetae flattened, widened, faintly serrated on both sided, distally narrowing into pointed tip ( Fig. 8C, D, F View FIGURE 8 ).
Two white lines running through ventral side of neuropodia and ventrum of each segment (except for tentacular segment) very indistinct, but detectable. Nephridial papillae on ventrum of segments 10, 11 and 12 prominent ( Fig. 7E View FIGURE 7 ), globular, inconspicuous in other segments, present from segment 5. Reduced parapodia of segment 18 lateral to pygidium, consisting of notopodia only. Pygidium conical. Anal cirri not observed.
Remarks. Macellicephala monroi sp. nov. can be distinguished from other members of the genus by a combination of the following characters: moderately developed, tapering prostomial lobes ( Fig. 2C, D View FIGURE 2 ), facial tubercle in form of an indistinct, enlarged pad, small to inconspicuous dorsal tubercules. Its distinct and rather striking pigmentation of purple and rusty brown horizontal stripes across dorsum can easily distinguish this species from other known Macellicephala .
Historic specimen of Monro (1930) ( Figs. 2C View FIGURE 2 and 7C View FIGURE 7 ) is in poor state with style of median antenna, tentacular and dorsal cirri all missing. All chaetae are now also missing, but at least some were observed by Monro (1930) as re-drawn here ( Fig. 8E, F View FIGURE 8 ). However, the characteristic purple and rusty brown pigmentation can be still clearly seen in this specimen ( Fig. 7C View FIGURE 7 ). Recently collected specimens ( Fig. 7A View FIGURE 7 ) from South Georgia as part of BIOPEARL I expedition were found to be morphologically consistent with historic material of Monro (1930). The best available specimen from the recently collected material was selected as holotype.
Additionally, two historic specimens from South Georgia, BMNH 1936: 2: 8: 488 were reported by Monro (1936) and later re-examined by Pettibone (1976). These specimens were considered Macellicephala mirabilis by both Monro (1936) and Pettibone (1976), while Monro also considered his previous record from 1930 of Macellicephala sp. (considered here M. monroi sp. nov.) to possibly represent the same species. Both specimens, BMNH 1936: 2: 8: 488 were re-examined here. The small specimen is in poor state, while the larger specimen may belong to another as yet undescribed species, but better-preserved material would be needed to establish that.
Molecular Information. Small, slightly damaged, but otherwise morphologically consistent specimen collected from Powell Basin, during BIOPEARL I expedition was chosen as a DNA voucher NHMUK.2018.75, with 16S (447 bp) and 18S (1161 bp) genes obtained from this specimen. The K2P distance to its sister species in the tree, Macellicephala brenesorum sp. nov., is 0.19 and uncorrected 'p' is 0.17, both values calculated from 16S as sequencing of COI was unsuccessful.
Etymology: This species is dedicated to Charles Monro for his contribution to taxonomy of polychaetes from
the Southern Ocean in the first half of 20th century, who also examined the historic material on which this description is partly based.
Distribution. Scotia Sea: South Georgia and Powell Basin, in depths of 230– 504m.



| NHMUK |
Natural History Museum, London |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Macellicephalinae |
|
Genus |
Macellicephala monroi
| Neal, Lenka, Brasier, Madeleine J. & Wiklund, Helena 2018 |
Macellicephala
| Monro (1930) : 47 |
Macellicephala mirabilis in Monro (1936) : 100
| Monro (1936) : 100 |