
Macellicephala gloveri, Neal & Brasier & Wiklund, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4455.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:70E92EDF-E11B-40A7-9573-0AC9F10E623E |
|
DOI |
https://doi.org/10.5281/zenodo.5994487 |
|
persistent identifier |
https://treatment.plazi.org/id/03BDA104-A330-6944-FF64-FF3AFABF23D7 |
|
treatment provided by |
Plazi |
|
scientific name |
Macellicephala gloveri |
| status |
sp. nov. |
Macellicephala gloveri View in CoL sp. nov.
( Figure 2G View FIGURE 2 , Figures 13 View FIGURE 13 , 14 View FIGURE 14 , and Figure 15A, B, C View FIGURE 15 ; Table 2.)
Macellicephala sp. in Brasier et al. (2016) [record].
Macellicephala sp. A in Neal et al. (2017) [record].
Material examined. Holotype, NHMUK.2018.211, Amundsen Sea , Southern Ocean, cruise JR 179, station BIO6 View Materials - EBS-3A, collected on 12/03/2008, epibenthic sledge, epi-net, 71°20'51.396''S, 110°0'47.808''W, 478 m depth GoogleMaps . Paratype (SEM specimen, on stub): NHMUK.2018.1007, Amundsen Sea , Southern Ocean, cruise JR 179, station BIO4 View Materials -EBS-3D, collected on 07/03/2008, epibenthic sledge, epi-net, 74°23'25.836''S, 104°46'4.512''W, 506 m depth. DNA voucher specimens (9 specimens) GoogleMaps : NHMUK.2018.855, Amundsen Sea , Southern Ocean, cruise JR 179, station BIO5 View Materials -EBS-1A, collected on 09/03/2008, epibenthic sledge, supra-net, 74°7'3.252''S, 105°50'31.272''W, 1472 m depth (1specimen) GoogleMaps ; NHMUK.2018.1463, Amundsen Sea , Southern Ocean, cruise JR 179, station BIO3 View Materials -EBS-1A, collected on 04/03/2008, epibenthic sledge, epi-net, 71°47'42.144''S, 106°13'9.588''W, 500 m depth (1 specimen) GoogleMaps ; NHMUK.2018.1422, Amundsen Sea , Southern Ocean, cruise JR 179, station BIO3 View Materials - EBS-1B, collected on 04/03/2008, epibenthic sledge, epi-net, 71°47'30.912''S, 106°12' 49.464''W, 500 m depth (1 specimen) GoogleMaps ; NHMUK.2018.9352 and NHMUK.2018.9353, Amundsen Sea , Southern Ocean, cruise JR 179, station BIO5 View Materials -EBS-2A, collected on 09/03/2008, epibenthic sledge epi-net, 73°52'46.704''S, 106°19'6.456''W, 1052 m depth (2 specimens) GoogleMaps ; NHMUK.2018.533 and NHMUK.2018.522, Amundsen Sea , Southern Ocean, cruise JR 179, station BIO5 View Materials -EBS-3D, collected on 10/03/2008, epibenthic sledge, epi-net, 73°58'40.152''S, 107°25'0.372''W, 550 m depth (2 specimens) GoogleMaps ; NHMUK.2018.9351 and NHMUK.2018.9356 Amundsen Sea , Southern Ocean, cruise JR 179, station BIO6 View Materials -EBS-2A, collected on 12/03/2008, epibenthic sledge, epi-net, 71°10'29.82''S, 109°51' 14.94''W, 1020 m depth (2 specimens) GoogleMaps .
Additional material examined: Macellicephala aciculata , holotype USNM 17405 About USNM , Pacific Ocean , Southern California, Albatross station 4352, collected on 14/03/1904.
Description (based on holotype and paratype). Medium sized species; holotype with 18 segments (segment 1 = tentacular segment), 13.5 mm long, 2 mm wide excluding parapodia and 5 mm wide including parapodia. Paratype (SEM specimen, on stub) with 18 segments, 13 mm long and 4.8 mm wide including parapodia. Live specimen ( Fig. 13A View FIGURE 13 ) pink with white prostomium and pink semi-circles present at anterior margin of each prostomial lobe; preserved specimen creamy yellow in colour ( Fig. 13B, C View FIGURE 13 ).
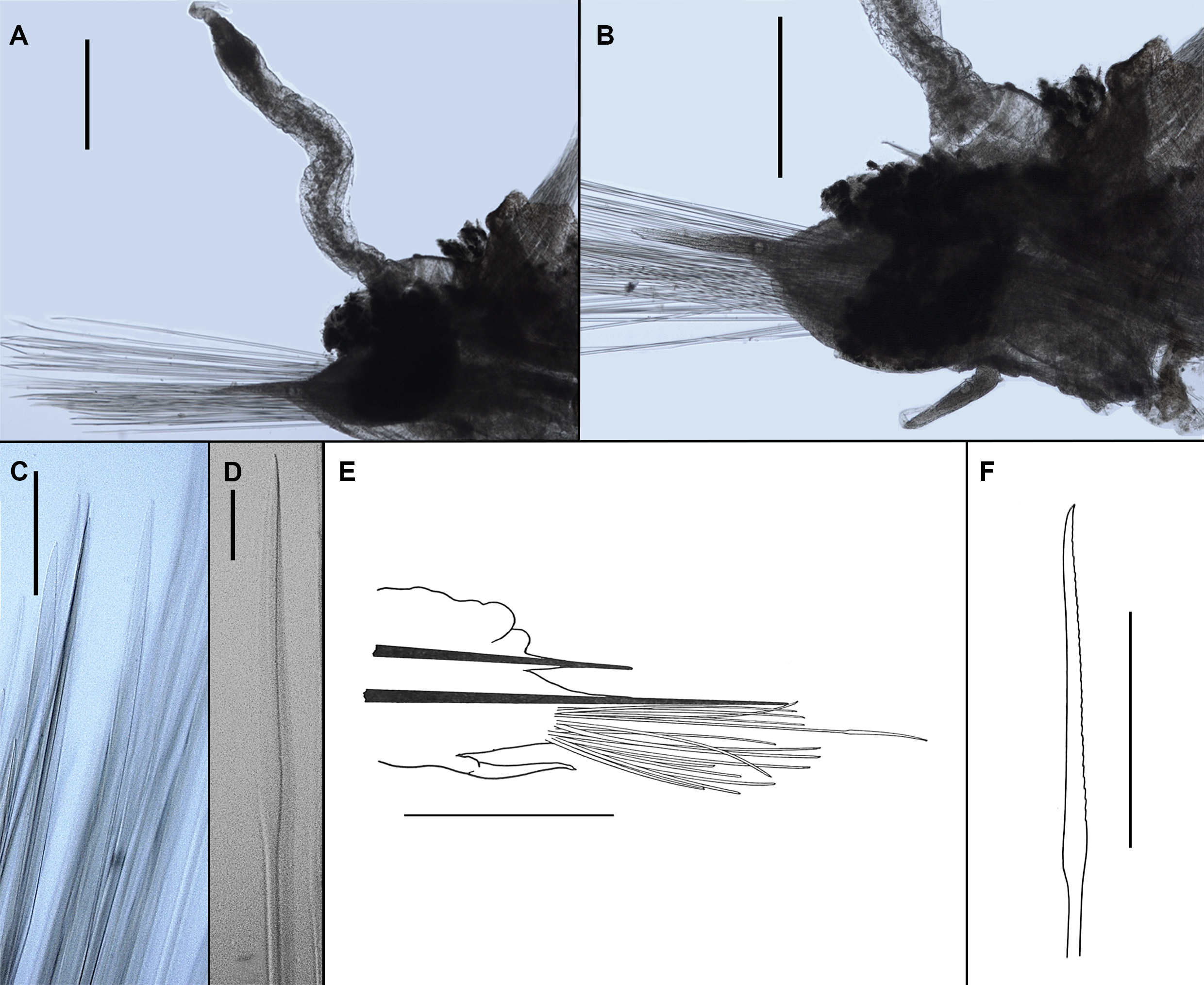
Prostomium bi-lobed, with deep anterior notch; prostomial lobes short (not extended), anteriorly rounded ( Figs. 2G View FIGURE 2 and 13D, E View FIGURE 13 ). Frontal filaments absent. Eyes absent. Ceratophore of median antenna large, inserted in anterior notch ( Fig. 13D, E View FIGURE 13 , Fig. 2G View FIGURE 2 ); style of median antenna missing. Palps very long (extending to segment 9), slender, smooth, tapering into slender tips ( Fig. 13A, B View FIGURE 13 ).
Tentaculophores inserted laterally to prostomium, both of equal size, achaetous ( Figs. 2G View FIGURE 2 and 13D, E View FIGURE 13 ); tentacular styles missing in type specimens. Facial tubercle absent. Everted pharynx with 2 pairs of jaws and 9 pairs of small triangular papillae of equal size ( Fig. 13C View FIGURE 13 ); each jaw with single fang and smooth margin.
Nine pairs of knob-like elytrophores present on segments 2, 4, 5, 7, 9, 11, 13, 15 and 17; elytra missing in type specimens.
Cirrigerous segments without dorsal tubercles; distinct dorsal cirrophores elongated, cylindrical to slightly bulbous; styles of dorsal cirri missing. Dorsal ridges appear faintly from segment 3 becoming obvious by segments 5-6, on segments 8-10 forming an oval structure, in segments anterior to segment 8 and posterior to segment 10 forming concentric structures ( Fig. 13C, F View FIGURE 13 ). In posterior part of body only, similar ridges appear on parapodia of cirrigerous segments; first parapodial ridge lateral to dorsal ridge, second parapodial ridge near base of cirrophore. All ridges appear ciliated and white in colour under light microscope but SEM revealed no ciliation of these structures ( Fig. 13F View FIGURE 13 ).
Parapodia biramous. Notopodia greatly reduced to one stout acicula ( Fig. 14A View FIGURE 14 ). Neuropodia large ( Fig. 14A View FIGURE 14 ), elongated, subtriangular to rectangular in shape, with integument covered acicula protruding into slender tip; prechaetal and postchaetal lobes poorly developed. Ventral cirrus inserted basally on segment 2, where very slender and long, approaching tip of neuropodial acicula lobe; inserted medially on subsequent segments where short (much shorter than tip of neuropodial lobe) ( Fig. 14A View FIGURE 14 ).
Notochaetae nearly as stout as neurochaetae; often entirely broken off (observed on only 7 parapodia in holotype), few (about 6 per ramus observed in holotype); very long (exceeding the length of neuropodial acicular lobe) ( Fig. 14A View FIGURE 14 ); with smooth, translucent basal half of shaft ( Fig. 14C View FIGURE 14 ); with obvious rose-bush like alternating and interlocking spines on one side only in distal half of shaft ( Fig. 14D View FIGURE 14 ), distal tip of notochaetae smooth and blunt ( Fig. 14 E, F View FIGURE 14 ). Neurochaetae short to very long, numerous, translucent, of two forms: either wide and flattened with shallow denticles on both sides throughout most of their lengths ( Fig. 14B View FIGURE 14 ) or slender, with rounded tip, with distinct rows of dense alternating denticles on both sides (few present and easily overlooked) ( Fig. 14B View FIGURE 14 , marked by arrow).
Nephridial papillae globular, present at ventral junction of neuropodia and body on segments 10, 11 and 12, inconspicuous in other segments, present from segment 5. Reduced parapodia of segment 18 lateral to pygidium, consisting of notopodia only. Pygidium rounded. Anal cirri not observed.
Additional morphological observation: Macellicephala gloveri sp. nov. was represented by 11 specimens in total. While DNA was successfully extracted from all specimens, confirming their identity, their morphological state was often rather poor. For example, the style of median antenna and dorsal cirri are missing in all available specimens. Tentacular styles are missing in type specimens but remained attached in three DNA vouchers: NHMUK.2018.9352 and NHMUK.2018.1463 (on right side only, in both specimens) and NHMUK.2018.855 (on left side only). When the tentacular cirri were observed these were smooth, very slender, both of equal length to palps. Length of palps appears to be variable as these extend to segment 9 in holotype, but are shorter (extending to segment 5) in most DNA voucher specimens, appearing shortest (extending to segment 3) in only 8 mm long DNA voucher specimen NHMUK.2018.9352. Finally, in DNA voucher specimen NHMUK.2018.855, single very small and easily overlooked elytra remained attached on segment 5 (left side only) ( Fig. 8C View FIGURE 8 ). Elytra are translucent, with smooth margins and sparsely covered by few microtubercules when observed under light microscopy.
Remarks. Semi-circular to circular dorsal and parapodial ridges described in Macellicephala gloveri sp. nov. and a closely related species from the Amundsen Sea M. linseae sp. nov., can also be also found in M. aciculata Moore, 1910 . Pettibone (1976) did not re-describe this character in detail; hence a type specimen of M. aciculata (USNM #1740) was examined, although it is in a very bad condition. The neuropodia of M. aciculata are oblique, tapering into slender tip with protruding acicula medially forming prechaetal and postchatel lobes. The neuropodia of M. gloveri sp. nov. are truncated, almost rectangular, with poorly developed prechaetal and postchaetal lobes. The stout notochaetae of M. aciculata appear to possess spines on both sides but in M. gloveri sp. nov. the alternating spines are confined to one side of the notochaeta only.
Based on morphological examination alone, the specimens from the Amundsen Sea were assumed to belong to one species. Subsequent molecular work [see Brasier et al. (2016)] revealed the presence of two distinct clades, suggesting presence of two species. Upon closer re-examination, some chaetal differences were observed in representatives of the two clades leading to erection of a second species as Macellicepala linseae sp. nov. The chaetal differences are as follows (see comparative Fig. 15 View FIGURE 15 ): notochaetae in M. gloveri sp. nov. shaft is smooth, translucent while in M. linseae sp. nov. it is straw-coloured and distinctly horizontally striated ( Fig. 15B and E View FIGURE 15 ). Further, notochaetae appear to be more numerous (~6), relatively slender and very long in M. gloveri sp. nov. ( Fig. 15A View FIGURE 15 ), while in M. linseae sp. nov. these are stouter, short and fewer in numbers (usually only one or two observed) ( Fig. 15D View FIGURE 15 ). Both species have two forms of neurochaete. The numerous, wide and flattened chaetae are very similar in both species. However, the additional few, slender neurochaetae appear to be different. In M. gloveri sp. nov. the serration is more dense, while in M. linseae sp. nov. the serration is more widely spaced (see Fig. 15C and F View FIGURE 15 ). As with notochaetae, neurochaetae are more numerous in M. gloveri sp. nov. Unfortunately, chaetae are susceptible to damage in both species and notochaetae in particular are often completely broken off, complicating the differentiation between these species. The differences observed are subtle and require careful observation but can be detected when specimens in good condition are available. It is possible that additional characters may be derived from elytra, which have unfortunately not been preserved in any of the specimens of M. linseae sp. nov. examined and only single observation of elytra was made for specimen of M. gloveri sp. nov.
Molecular Information. COI and 16S were sequenced by Brasier et al. (2016), while 18S (1731 bp) was obtained in this study ( Table 1). Based on COI, this species was closest to Macellicephala linseae sp. nov. with K2P distance 0.12 and uncorrected 'p' distance 0.11.
Etymology. This species is named after Dr. Adrian Glover from the Natural History Museum, London who collected the polychaetes during the BIOPEARL II project and managed subsequent work on the polychaete material [see Neal et al. (2012); Neal et al. (2014); Brasier et al. (2016); Neal et al. (2017)].
Distribution. Amundsen Sea, including Pine Island Bay at shelf and slope depths of 500–1072 m.




| NHMUK |
Natural History Museum, London |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Macellicephalinae |
|
Genus |