
Rhynchelmis orientalis Yamaguchi, 1936
|
publication ID |
https://doi.org/ 10.5281/zenodo.194252 |
|
DOI |
https://doi.org/10.5281/zenodo.6201853 |
|
persistent identifier |
https://treatment.plazi.org/id/03C5878E-FFE9-D418-B2CF-F8CA7B6D6A1C |
|
treatment provided by |
Plazi |
|
scientific name |
Rhynchelmis orientalis Yamaguchi, 1936 |
| status |
|
Rhynchelmis orientalis Yamaguchi, 1936 View in CoL
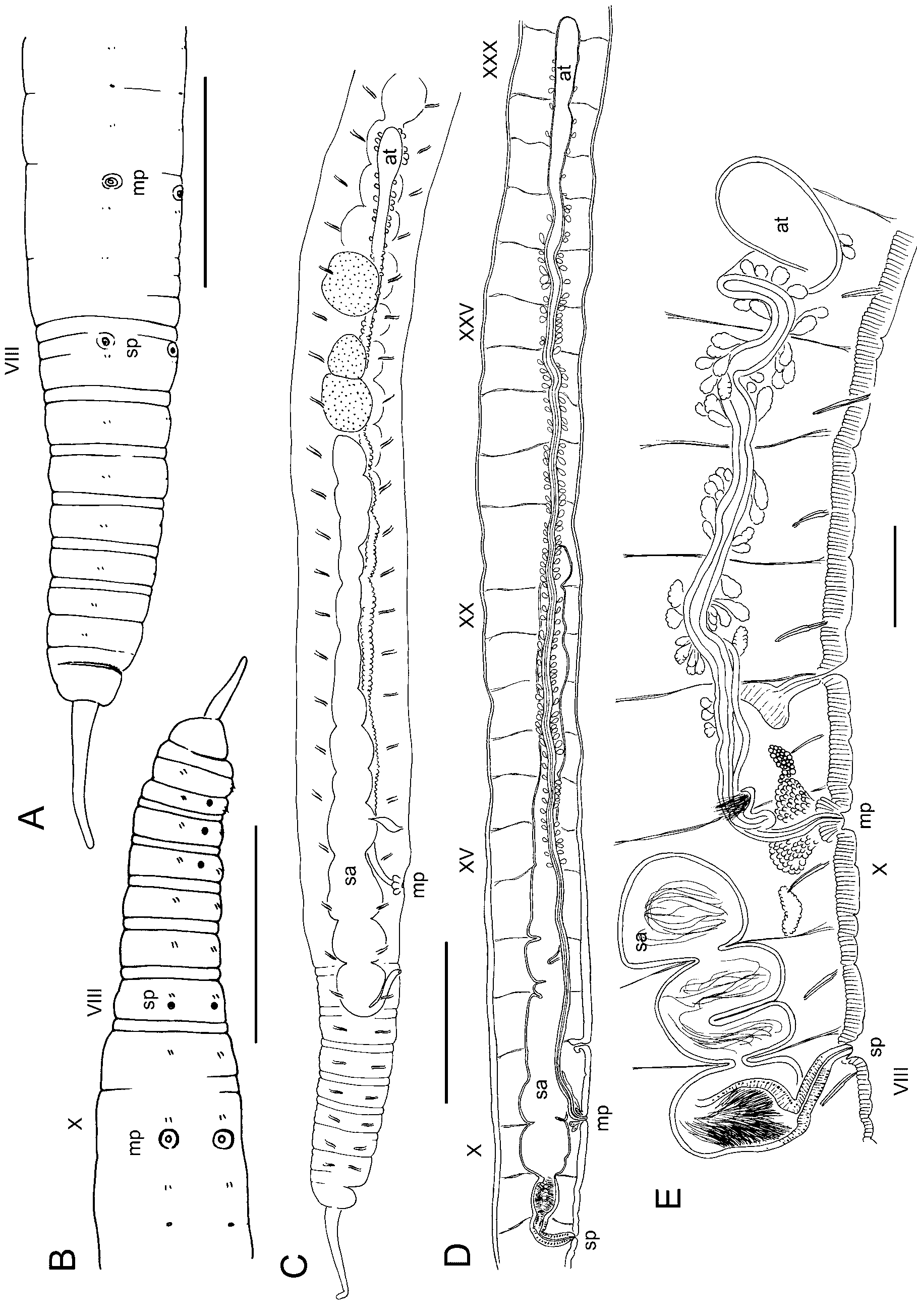
( Figs. 6–8 View FIGURE 6. R View FIGURE 7. R View FIGURE 8. R )
Supplementary description. Material from the type locality: Body diameter 1.3–1.4 mm in X, length about 70 mm, 121–125 segments. Annulation moderately developed, but obscured in clitellum ( Figs. 6 View FIGURE 6. R A–B); body round in transverse section, and dorsolateral transverse muscles weak. Clitellum from IX to XVI or XVIII. One slightly post-mature specimen with row of midventral pores in III–V, 70 Μm wide ( Fig. 6 View FIGURE 6. R B). Midventral pores associated with bundles of cell extensions from several clusters of gland cells. Gland cell bundles 100– 200 Μm long, usually 2 on either side of ventral nerve cord, and 1 or 2 beneath it ( Fig. 7 View FIGURE 7. R A). No midventral glands on other mature or nearly-mature worms.
Longitudinal muscle bands without tapered lateral edges; edges do not curl into coelomic space ( Figs. 7 View FIGURE 7. R B, 8B). Pharynx roof thick in II, but does not form distinct dorsal pad; pharyngeal glands IV–VII. Circulatory system as originally described ( Yamaguchi 1936): starting near X, two pairs of lateral vessels have up to 5 or 6 long distal branches; one branch of the anterior pair joining the perivisceral sinus. Posterior to about XL, main branches of both lateral vessels join the perivisceral sinus ventrolaterally. Few short, blind branches (caecae) on dorsal part of lateral vessels in anterior segments, but number increases posteriorly, up to 10 or more posterior to about XL.
Nephropores simple, without vesicle, just anterior to ventrolateral chaetae. First nephridia on 12/13, but tubule may extend forward a segment or two. A strong dorsal tubule loop, as in other Rhynchelmis ( Fig. 7 View FIGURE 7. R C) extends posteriorly to the midventral region. Nephridia paired in most mid-body segments.
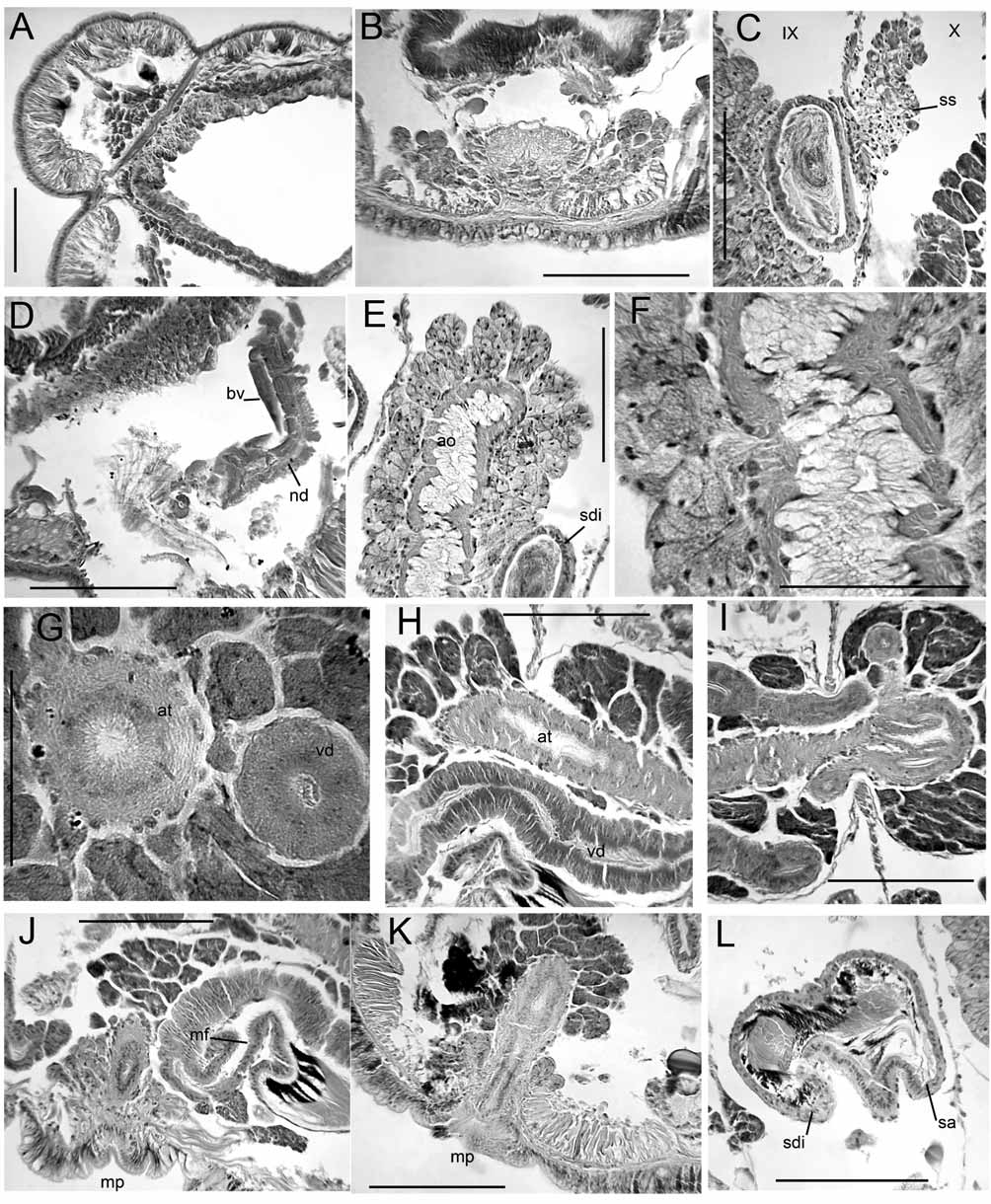
Spermathecal pores without epidermal modification or distinct glands ( Figs. 7 View FIGURE 7. R B, D). Spermathecae very long, passing through 8/9 and 9/10, then entering the sperm sac for several segments, to XXI in one mature worm ( Fig. 6 View FIGURE 6. R D). Spermathecae with cylindrical ectal duct, about 500 Μm long by 110 Μm wide, followed by a long, tubular ampulla without diverticula or gut connection, distended between septa up to 600 Μm wide in mated worms ( Figs. 6 View FIGURE 6. R C–D). Ectal duct with columnar epithelium having basal nuclei; ampulla with cuboidal epithelial cells; muscle layer less than 5 Μm thick ( Figs. 7 View FIGURE 7. R D–H). Ectal end of ampulla (in VIII or IX) histologically distinct, with epithelium having distinct nuclei ( Fig. 7 View FIGURE 7. R E); entally, the epithelium with nuclei small or absent, and with granular vacuoles ( Figs. 7 View FIGURE 7. R G–H). Heads of sperm cells lined up around periphery of anterior portion of ampulla in mated worm ( Fig. 7 View FIGURE 7. R E); loose sperm in remainder ( Fig. 7 View FIGURE 7. R F).
Male pore without penis or penial bulb, surrounded by 200 Μm wide circle of thickened epidermis, internally surrounded by a low mass of prostate-like, granular glands to 60 Μm high ( Figs. 8 View FIGURE 8. R A–B). Atria to XXXI and XXXIII in the two newer specimens (total atrium length to nearly 14 mm); to XXII in the original specimen. Main portion of atrium a narrow tube, diameter 60–70 Μm, but entally expanded to as much as 300 Μm ( Figs. 6 View FIGURE 6. R C–D, 8F). Muscle layer thin, 2–5 Μm throughout. Multicellular prostate glands relatively sparse ( Figs. 8 View FIGURE 8. R C–H), especially near ental end ( Fig. 8 View FIGURE 8. R F); atrium without prostates in ectal portion, from X to XI or XIII ( Fig. 8 View FIGURE 8. R C). Posterior vas deferens diameter up to 20–30 Μm; vas does not penetrate 10/11, enters muscle layer of atrium near beginning of prostates ( Figs. 8 View FIGURE 8. R C–D), and runs inside the atrial wall to near ental end ( Figs. 8 View FIGURE 8. R E,G). Anterior vas deferens diameter less than 10 Μm, degenerates in X in mature worms, but traces may persist in ental portion, mostly embedded in atrial muscle or adjacent to atrium. Single pair of testes in X, and ovaries in XI. Sperm sacs to XLVI in one mature worm, to LXXXI in the other; egg sacs with ova to LVI and XCIV.
Material from the Okoppe River: Body diameter 0.68–0.82 mm in X, tails missing on both mature worms; one complete immature worm with 127 segments. One mature specimen with midventral glands in III–V, the other with a single gland in V. Circulatory system generally less-developed than in material from the type locality. Anterior pair of lateral blood vessels from X to mid-body may be unbranched, or with few branches; posterior pair may be very short. Lateral blood vessels in posterior segments (beginning about LX or LXX) with more extensive branching, and with several short, blind caecae in the dorsal part.
Spermathecae extend posteriorly to X in one mature worm, but as far as XV in the other. Ectal duct of spermatheca about 220–290 Μm long by 52–58 Μm wide, ampulla up to 190 Μm wide, histologically differentiated as in material from the type locality ( Fig.6 View FIGURE 6. R E). Two partially-mature worms with large cell masses (possibly developing spermathecal sacs) extending posteriorly from 8/9 and 9/10. Male pore within a 90 Μm wide circle of non-clitellar epidermis, internally surrounded by a low mass of petiolate gland cells to 150 Μm high ( Fig. 6 View FIGURE 6. R E). Atria to XV and XXVI in the two mature specimens. Diameter of ectal portion 30–60 Μm; ental expansion to 180–240 Μm; total atrium length 2200–5300 μm. Posterior vas deferens diameter up to 19 Μm, joining atrium before beginning of prostates; posterior sperm funnel well developed. Anterior vas deferens diameter up to 10 Μm, without anterior sperm funnel or with a vestigial funnel on one worm. Sperm sacs to XXV in one mature worm, to XXXIV in the other; egg sacs with ova to XXXII and XXX.
Remarks: R. orientalis
Only a single specimen of R. orientalis apparently remains from the original series (Dr. A. Ohtaka, personal communication). This specimen was apparently not quite mature, as the clitellum was not distinct and no sperm was present in the spermathecae. Of the three additional specimens from the type locality, one was mature, with spermathecae and atria filled with sperm; another was apparently somewhat post-mature, with small testes and sperm tails in spermathecae and atria. The latter specimen had a row of midventral pores in III–V ( Fig. 6 View FIGURE 6. R B), associated with bundles of cell extensions from several clusters of gland cells ( Fig. 7 View FIGURE 7. R A). Mature specimens from the Okoppe River had similar midventral (subneural) glands; both of these worms had sperm in the spermathecae and mature eggs. The number of glands varied in the latter specimens, suggesting that this character is variably expressed. Yamaguchi (1936) did not observe these glands in the type series of R. orientalis , although mature material may have been limited. The new collection of Rhynchelmis from the Okoppe River indicates that the species is not limited to Hokkaido, but also occurs in at least the northern part of Honshu. The relatively small size, shorter atria and reduced blood vessels suggest variation among populations of this species, similar to that observed in some Nearctic species ( Fend & Brinkhurst 2000).
Cook (1971) examined the orientalis type material, and considered this species to be very close to the Western Nearctic R. elrodi , for which the genus Rhynchelmoides ( Hrabĕ 1936) was erected. Other, mostly western Nearctic species have been associated with this group, either formally as comprising the genus Rhynchelmoides (e.g., Hrabĕ 1982, Sokolskaya 1983) or informally ( Fend & Brinkhurst 2000, Holmquist 1976). These include R. glandula , R. alaskana , Rhynchelmis saxosa Fend & Brinkhurst, 2000 , R. brooksi and R. malevici . Hrabĕ (1982) considered Rhynchelmoides to be Nearctic, but did not mention the Palearctic R. orientalis ; R. malevici and R. saxosa had not yet been described. Both Hrabĕ (1982) and Sokolskaya (1983) included R. brooksi in Rhynchelmoides, but ignored the Nearctic R. rostrata (Eisen) , which had originally been described under the genus Sutroa , but later combined with Rhynchelmis ( Brinkhurst & Cook 1966) . The analytical results of Kaygorodova & Liventseva (2007) grouped elrodi , glandula and orientalis , together with the Sutroa group species in a separate clade from alaskana , saxosa , brooksi and R. malevici , which in turn grouped with the R. ( Rhynchelmis ) species. Thus the species that traditionally have been associated with Rhynchelmoides were not resolved as a clade in that analysis (their Fig. 2 View FIGURE 2. R ), and orientalis did not group with the other species lacking a spermatheca-gut connection. The discrepancies between the latter, analytical results and the earlier, intuitive classification based on overall similarity relates to the lack of obvious synapomorphies for the Rhynchelmoides group, compared with the other species groups within Rhynchelmis .
Although the earlier definition of Rhynchelmoides was quite limited ( Hrabĕ 1936), an expanded set of characters was given by Hrabĕ (1982): the absence of R. ( Rhynchelmis )-like curled muscles, the cylindrical body, the absence of anterior vasa deferentia and male funnels, the vasa deferentia entering the atrial wall near the ectal end and running under the muscle layer to near the ental end, and the narrow diameter of the vasa deferentia compared with the atria. The absence of curled muscles (assumed by their omission in the literature description) was used by Sokolskaya (1983) to associate R. orientalis with Rhynchelmoides. The new material confirms that the body is round, or at least not strongly quadrilateral, and longitudinal muscles are not tapered or curled. In addition, septal grooves are not as strong as in the R. ( Rhynchelmis ) species, and the septa are obscured in the clitellum ( Figs. 6 View FIGURE 6. R A–B).
Rhynchelmis orientalis View in CoL differs from other Rhynchelmoides group species in having a vestigial anterior vas deferens, and can be interpreted as resembling the Sutroa View in CoL group in this respect. The condition of the anterior vas actually seems intermediate between the Sutroa View in CoL and Rhynchelmoides group character states, as it is absent or weakly expressed in the atrial muscle posteriorly. Thus the loss of anterior male ducts (see Brinkhurst 1989) may still be argued to be a potential synapomorphy for the Rhynchelmoides group (including R. orientalis View in CoL ). Unlike other Rhynchelmoides group species, the vasa deferentia do not immediately embed in the atrial wall near the male pore, and the beginning of the prostate layer is well posterior to X. However, the embedded vas and the prostate layer do extend over most of the atrial length. Yamaguchi (1936) described atria and sperm sacs extending as far back as XXVII in one specimen, which is farther than in any other Rhynchelmis View in CoL species; in some of the new material the atria are even longer. The extent of the sperm and egg sacs in specimens from the type locality is also unusual.
As apparent symplesiomorphies, the body wall characters do not resolve the Rhynchelmoides group as a monophyly. A few additional similarities, whether apomorphic or plesiomorphic, may be significant in future morphological analyses. The arrangement of blood vessels (two branched pairs per segment) of R. orientalis View in CoL resembles that of most Nearctic species in the Sutroa View in CoL and Rhynchelmoides groups. The general form and histology of the atrium resembles that of most other Rhynchelmoides group species (except brooksi View in CoL and malevici View in CoL ), and lacks the complex musculature of most Sutroa View in CoL group species. As in the case of R. saxosa View in CoL , R. alaskana View in CoL , and to a lesser extent R. elrodi View in CoL and R. glandula View in CoL , the ental end of the atrium is strongly expanded; as with R. saxosa View in CoL , the prostates may be sparse entally. Although it is relatively long, the R. orientalis View in CoL spermatheca is similar to those of the other Rhynchelmoides group species lacking a gut connection: the ectal end of the ampulla has concentrated sperm and the ental end is lined with unmodified epithelium. The midventral glands in anterior segments of some specimens are unusual in the family, and are a likely synapomorphy with R. glandula View in CoL , R. alaskana View in CoL , and R. saxosa View in CoL .
Unlike other Rhynchelmoides group species (plus R. brooksi and R. malevici ), the ectal duct of the spermatheca is not thickened. Male and spermathecal pores are simple, without obvious penes, in contrast to the small but distinctive penes of some Rhynchelmoides group species. Although Yamaguchi described the first nephridia as being in XIV, they are on 12/13, with pores in XIII in our material, as in other members of the genus; the dorsal loop of the nephridial tubules is also typical for Rhynchelmis .


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.