
Talorchestia spinipalma ( Dana, 1852a )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4319.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:67558747-9346-4561-8Ca5-Fb15Cdc7840B |
|
DOI |
https://doi.org/10.5281/zenodo.6021812 |
|
persistent identifier |
https://treatment.plazi.org/id/03CF87EB-FFF9-F14F-CB8A-9B06FC15C6F9 |
|
treatment provided by |
Plazi |
|
scientific name |
Talorchestia spinipalma ( Dana, 1852a ) |
| status |
|
Talorchestia spinipalma ( Dana, 1852a) View in CoL
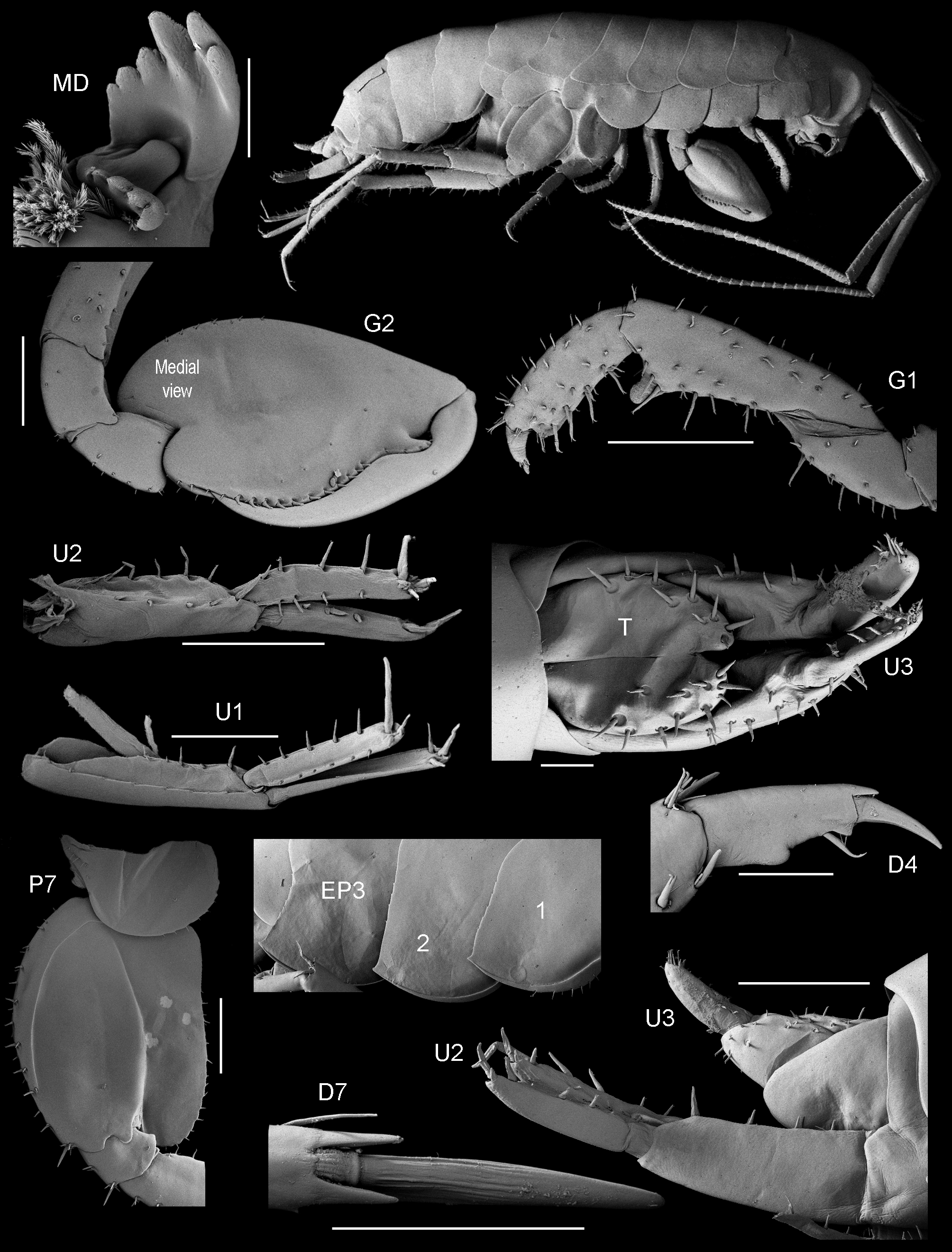
( Fig. 18 View FIGURE 18 )
Orchestia spinipalma Dana, 1852a: 203 View in CoL .— Dana, 1853: 875, pl. 59, fig. 4a–e.— Spence Bate, 1862: 28, pl. 4, fig. 9. Talorchestia spinipalma View in CoL . — Stebbing, 1906: 552 (in part, part = T. terraereginae View in CoL ).— Stephensen, 1935: 12 (no new records).—? Schellenberg, 1938: 66.— J.L. Barnard, 1960: 24, figs 7, 8).—? Bousfield, 1970: 163.— Morino & Miyamoto, 1988: 95, figs 4–6. — Lowry & Stoddart, 2003: 276 (including T. terraereginae Haswell, 1880 View in CoL ).— Lowry & Springthorpe, 2009: 905.— Serejo, 2009: 897, figs 3, 4.— Lowry & Bopiah, 2013: 354, figs 5–8.
Type material. Neotype, male, 16.5 mm, AM P.87317, fine white sand on steep narrow beach just north of Liku’alofa Beach Resort , Tongatapu, Tonga (21°04.838'S 175°20.652'W), white talitrids under dried Turbinaria and other algae, J.K. Lowry & G.C.B. Poore, 14 October 2011. GoogleMaps
Type locality. Fine white sand on steep narrow beach just north of Liku’alofa Beach Resort, Tongatapu, Tonga (21°04.838'S 175°20.652'W). GoogleMaps
Material examined. 9 specimens, AM P.75984; 1 male, AM P.99072 (SEM pin and 4 SEM stubs); 1 female AM P.99073 (wet specimen and SEM stub), Mololo Beach, Madang, Papua New Guinea (4°58'S 145°47'E) supralittoral sand, J.D. Thomas, 4 February 1990 GoogleMaps .
Habitat. White talitrids under wrack in the supralittoral zone.
Diagnostic description. Antenna 2 more than half but not exceeding body length (about 90¾). Gnathopod 1 carpus 1.6–1.7 × as long as propodus, about 3.3 × as long as broad; propodus tapering. Gnathopod 2 merus and carpus fused; propodus about 2.5 × as long as wide, palm reaching about 65¾ along posterior margin, with strong rounded protuberance near dactylar hinge. Epimeron 1 with 7 robust setae along ventral margin. Epimeron 3 with small subacute tooth. Uropod 2 inner ramus subequal in length to outer ramus. Uropod 3 ramus 2.9 × as long as broad. Telson with about 8–14 setae per lobe.
Remarks. There is significant morphological variation between the geographical populations of T. spinipalma (table 1). The populations from Mololo beach, Madang, Papua New Guinea and the neotype described by Lowry & Bopiah (2013) from Tonga are most similar. The adult males are 11 mm and 16.5 mm respectively, the ramus of uropod 3 is slightly longer 2.9 mm in the Madang material, 2.6 mm in the Tongan material. The Madang material has more robust setae per lobe on the telson (9–11) that the Tongan material (7–8).
Neither Schellenberg 1938 (Ralum, Bismarck Archipelago and Bohol, Philippine Islands), nor Bousfield 1970 (Rennell Island, Solomon Islands), illustrated their specimens and so what species they had is problematic. It is difficult in general to know which specimens reported as T. spinipalma are that species. The specimen from Kosrae (as Kusaie), Micronesia, illustrated by J.L. Barnard (1960), was 13 mm in length with 10–13 robust setae on the telson and a relatively short palm on male gnathopod 2. It is similar to the 16.5 mm neotype of Lowry & Bopiah (2013) and the differences can be attributed to a smaller growth stage. The 10 mm specimen illustrated by Morino & Miyamoto (1988) had six robust setae on the peduncle of uropod 3, 11–13 robust setae on the telson and a relatively short palm to posterior margin length on male gnathopod 2. Serejo’s (2009) 13 mm specimen from the Queensland coast falls into the same category. The specimens reported here from Mololo beach, near Madang, Papua New Guinea are most similar to the specimen reported from Tongatapu, Tonga by Lowry & Bopiah (2013). The one thing all of the illustrated populations have in common, is a fused merus and carpus on male gnathopod 2. In the Tongan population of T. spinipalma the fusion appears to be between the carpus/propodus whereas in the Madang and Kosrae populations the fusion is clearly between the merus/carpus.
The Madang and Tonga populations of Talorchestia spinipalma are similar to Talorchestia mindorensis from the Philippines and Taiwan. Both have the strong palmar protuberence near the dactylar hinge on the male second gnathopod and the dactylus reaching about 65¾ along the posterior margin of the propodus. But T. mindorensis has a short second antenna (less than half the body length) and a shorter carpus on gnathopd1 (length about 2.6 × breadth), and significantly, the carpus not fused.
Distribution. Australia. Queensland, Port Douglas ( Serejo 2009). Marshall Islands. Yap; Kosrae (as Kusaie Island) ( J.L. Barnard 1960). New Caledonia. Nou Vata, Noumea; Ile of Pines ( Morino & Miyamoto 1988). Papua New Guinea.? Ralum, Bismarck Archipelago ( Schellenberg 1938); Madang (this study); Motupore Island ( Morino & Miyamoto 1988).? Philippine Islands. Ubay, Bohol ( Schellenberg 1938). Solomon Islands. Gizo ( Morino & Miyamoto 1988).? Rennell Island ( Bousfield 1970). Tonga. Tongatapu ( Dana 1852a, Lowry & Bopiah 2013).
Type material. Holotype, male, 16.5 mm, AM P.99068 (SEM pin and 4 SEM stubs), Kuta Beach , Central Lombok, Indonesia (8°53.667'S 116°17.050'E), supralittoral sand, Dwi Listyo Rahayu GoogleMaps . Paratypes: female (SEM pin and SEM stub), male “b” (SEM stub), 157 males and females, AM P.99069, same data as holotype GoogleMaps .
Type locality. Kuta Beach , Central Lombok, Indonesia (8°53.667'S 116°17.050'E). GoogleMaps
Ecological type. Beach-hoppers (mainly coastal supralittoral/intertidal leaf-litter/wrack, non-substrate modifying talitrids).
Etymology. The new species is named after Dr Dwi Listyo Rahayu (Lembaga Ilmu Pengetahuan (Indonesian Institute of Sciences), Pemenang, Lombok, Indonesia), known as Yoyo to her friends and colleagues, for her important contribution to South-East Asian carcinology and for making this new species available for study; used as a noun in apposition.
Description. Based on holotype, male, 16.5 mm, AM P.99068.
Head. Ventroproximal surface with 5 groups of long, slender scales. Antenna 2 more than half but not exceeding body length; article 5 longer than article 4; flagellum final article large, cone-shaped forming a virgula divina, with apical cluster of 'imbricated' setae. Labrum with apical setal patch; epistome without robust setae, without many pores. Mandible left lacinia mobilis with 6 cusps (sixth minute). Labium with vestigial inner plates. Maxilla 1 with small palp, 2-articulate.
Pereon. Gnathopod 1 coxa smaller than coxa 2; damaged, carpus damaged, about 3 × as long as broad. Gnathopod 2 basis slender; ischium without lobe on anterior margin, distal triangular small anterodistal lobe on medial surface; carpus and merus separate; carpus triangular, reduced (enclosed by merus and propodus), posterior lobe absent, not projecting between merus and propodus; propodus subovate, 1.8 × as long as wide, palm reaching about 77% along posterior margin, weakly toothed, incised or sinuous, with strong rounded protuberance near dactylar hinge, lined with (12 + 7) medial robust setae, without midpalmar protuberances, posteromedial surface of propodus with groove, without cuticular patch at corner of palm; dactylus subequal in length to palm, sinusoidal, without anteroproximal bump, posterior margin lined with small robust setae, with posteroproximal sinus and small projection. Pereopods 2–4 coxae wider than deep, or as wide as deep. Pereopods 3–7 dactyli with or without anterodistal patch of many rows of tiny setae (vestigial on pereopod 5). Pereopod 3 dactylus without anterodistal patch. Pereopod 4 significantly shorter than pereopod 3; carpus significantly shorter than carpus of pereopod 3; dactylus without anterodistal denticular patch. Pereopod 5 propodus distinctly longer than carpus; dactylus with vestigial anterodistal denticulate patch. Pereopod 6 coxa posterior lobe medial view posteroventral corner rounded, posterior margin oblique to ventral margin, posterior lobe without ridge, posterior lobe with 14 marginal setae. Pereopod 7 basis lateral sulcus present, slightly pronounced, posterodistal lobe absent; distal articles (merus and carpus) slender; merus posterior margin straight.
Pleon. Epimeron 1 with 3 robust setae along ventral margin. Epimeron 2 subequal in length to epimeron 3. Epimeron 3 posterior margin smooth, with setae (minute), posteroventral corner with small subacute tooth. Uropod 1 peduncle with 9 robust setae, distolateral robust seta small (less than 1/4 length of outer ramus); inner ramus slightly shorter than outer ramus, with 4 medial and 4 lateral robust setae; outer ramus without marginal robust setae. Uropod 2 peduncle with 8 robust setae; inner ramus with 3 medial and 3 lateral robust setae; outer ramus with 2 marginal robust setae. Uropod 3 peduncle with 7–8 robust setae; ramus partially fused to peduncle, 3.1 × as long as broad, linear (narrowing), with 4 marginal robust setae, with more than 5 apical robust setae. Telson dorsal midline complete, 8–10 setae per lobe.
Remarks. Talorchestia yoyoae belongs in the species group with a rounded protuberance on the male gnathopod 2 palm near the dactylar hinge. The gnathopod 2 dactylus is long, reaching more than 75¾ along the posterior margin of the propodus, similar to T. bunaken . Talorchestia yoyoae differs from T. bunaken as discussed in the remarks for T. bunaken .
Distribution. Indonesia. Kuta Beach, Central Lombok (this study).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Senticaudata |
|
Family |
|
|
Genus |
Talorchestia spinipalma ( Dana, 1852a )
| Azman, B. A. R. 2017 |
Orchestia spinipalma
| Lowry 2013: 354 |
| Lowry 2009: 905 |
| Serejo 2009: 897 |
| Lowry 2003: 276 |
| Morino 1988: 95 |
| Bousfield 1970: 163 |
| Barnard 1960: 24 |
| Schellenberg 1938: 66 |
| Stephensen 1935: 12 |
| Stebbing 1906: 552 |
| Spence 1862: 28 |
| Dana 1853: 875 |
| Dana 1852: 203 |