
Sabellaria salazari, Chávez-López, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5048.2.3 |
|
publication LSID |
lsid:zoobank.org:pub:CBD94592-5647-4F52-A8CF-4990DE8574D8 |
|
DOI |
https://doi.org/10.5281/zenodo.5555043 |
|
persistent identifier |
https://treatment.plazi.org/id/03D0AE23-5169-FFC5-FF73-FE51FCBB12A5 |
|
treatment provided by |
Plazi |
|
scientific name |
Sabellaria salazari |
| status |
sp. nov. |
Sabellaria salazari View in CoL n. sp.
Figures 9A– F View FIGURE 9 ; 10A–N View FIGURE 10
LSID: urn:lsid:zoobank.org:act:83477E2E-D671-4958-9495-325DB172B0A2
Material examined. Quintana Roo. Holotype (ECOSUR- 254), Cozumel, Playa Azul , on Millepora , February 26, 1999, coll. J. R. Bastida-Zavala & S.I. Salazar-Vallejo ). Campeche. Paratypes ECOSUR-255, five spec. ( Champotón , on rock, 4–5 m, February 16, 1999, coll. J. R. Bastida-Zavala & S.I. Salazar-Vallejo). ECOSUR-256, one spec. (Champotón, on rock, 3 m, February 15, 1999, coll. J. R. Bastida-Zavala & S.I. Salazar-Vallejo). ECOSUR- 257, nine spec. (Champotón, among seaweed and seagrasses, on the beach, February 14, 1999, coll. J. R. Bastida-Zavala & S.I. Salazar-Vallejo) .
Additional material examined. Yucatán: ECOSUR-P3205, three spec. (2 km off Río Lagartos , February 18, 1999, on coral rock, 2–4 m, coll. J. R. Bastida-Zavala & S.I. Salazar-Vallejo) . ECOSUR-P3206, one spec. ( San Felipe ; February 19, 1999, on coral rock, 2 m, coll. J. R. Bastida-Zavala & S.I. Salazar-Vallejo) . BCC-13351, one spec. ( Northeast of Progreso , 21.492633, -89.557483, 15.3 m, November 17, 2015, col. SBZ) GoogleMaps . Campeche. ECO-SUR-P3207, one spec. ( Términos Lagoon , July 1984) . Florida: UF, one spec. (Cedar Keys, Seahorse Key , Florida, March 31, 1973) . UF3457 , colony ( Seahorse Key , Florida, December 13, 2012, coll. Gustav Paulay et al.) . UF4596 , one spec. ( Seahorse Key , Florida, January 26, 2016, intertidal, 0–1 m, on sand flat) .
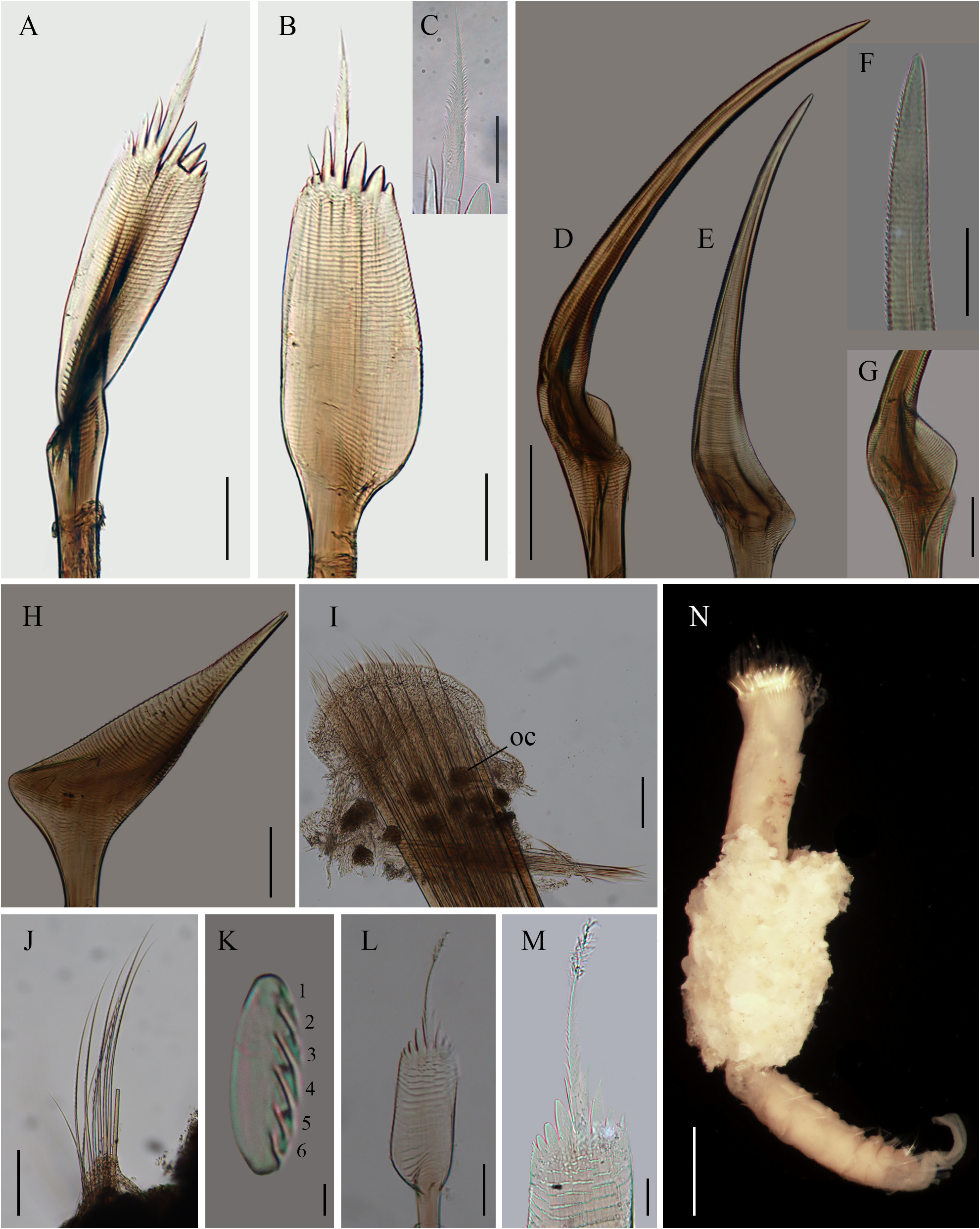
Description. Holotype (ECOSUR-0254), complete, mature female. Body colorless, 12.5 mm long, 1.5 mm wide, with three parathoracic segments, 23 abdominal segments, caudal peduncle 1.5 mm long ( Fig. 9 View FIGURE 9 ).
Opercular crown and opercular stalk partially fused ( Fig. 9A, D View FIGURE 9 ). Opercular disc oblique, three rows of amber paleae; outer row with 15–16 paleae per side, middle and inner rows each with 7–8 paleae per side. Outer paleae with three pairs of lateral teeth; blade flat, oblong, almost three times longer than wide; thecae transverse with finely serrate margins ( Fig. 10A–B View FIGURE 10 ). Lateral teeth wider on one side ( Fig. 10B View FIGURE 10 ). Medial spike straight, ½ as long as blade, smooth basally, distally penicillate along 2/3 total length ( Fig. 10C View FIGURE 10 ). Dorsal paleae, in lateral view, with the blade slightly twisted, bent towards outside of opercular crown; blade and handle at an angle of 160° to 170° ( Fig. 10A View FIGURE 10 ). Middle paleae geniculate, long, almost twice longer than outer paleae; blade in a convex angle of 125° to 140° in dorsal paleae to ventral paleae, transverse thecae with finely serrated margins ( Fig. 10D View FIGURE 10 ); tip blunt with finely serrate margin on one side ( Fig. 10F View FIGURE 10 ). Blade base concave with a lateral expansion ( Fig. 10G View FIGURE 10 ) decreasing in size from dorsal to ventral position ( Fig. 10D–E View FIGURE 10 ). Inner paleae geniculate, ½ as long as middle paleae; concave blade directed inwards, thecae transverse with finely serrate margin, tips blunt, smooth ( Fig. 10H View FIGURE 10 ).
Opercular peduncle colorless with some dark spots ( Fig. 9A–B View FIGURE 9 ). Three pairs of nuchal spines embedded in opercular tissue ( Fig. 9E View FIGURE 9 ). Tentacles and palps pale with brown speckles ( Fig. 9B–C View FIGURE 9 ). Median ridge as long as the opercular stalk, with dark marginal eyespots. Median organ broken. Building organ colorless.
Thorax without notochaetae. Lateral sides of thoracic segments with very dark pigment ( Fig. 9A–B View FIGURE 9 ). Chaetiger 1 with a pair of neuropodia bearing capillary chaetae. Chaetiger 2 with capillary neurochaetae and a pair of small cirri and branchiae.
Parathorax with three segments, all with paired branchiae. Lateral sides of first parathoracic segment intensely dark, notochaetae and neurochaetae dark amber. Second segment with paler pigmentation, chaetae amber. Third segment colorless, chaetae amber ( Fig. 9B View FIGURE 9 ). Notopodia with seven lanceolate and capillary chaetae. Neurochaetae lanceolate thinner than notochaetae ( Fig. 10I View FIGURE 10 ).
Abdominal segments colorless, with a pair of branchiae decreasing in size up to segment 11. Neuropodia with light brown dots from the sixth abdominal segment onwards. Neurochaetae verticillate ( Fig. 10J View FIGURE 10 ). Notopodia with a series of uncini with 5–6 transverse rows of teeth ( Fig. 10K View FIGURE 10 ). Caudal peduncle colorless, slightly annulate; distally round, flat, translucent.
Parathoracic and abdominal parapodia with oocytes, each about 70 µm in diameter ( Fig. 10I View FIGURE 10 ).
Variation. Body 1.4–12 mm in total length, 0.2–1.5 mm wide, with 6–23 abdominal segments, caudal peduncle 0.1–1.5 mm long. Specimens reaching 5 mm in total length showed speckled tentacles. Opercular crown made of 12–23 outer paleae per lobe, 6–13 middle paleae per lobe, and 6–12 inner paleae per lobe. Nuchal spines protruding from opercular tissue in small specimens (<6 mm total length).
Outer paleae with 3–5 pairs of lateral teeth, commonly with 3–4 larger teeth. Outer paleae of small specimens 2 mm in total length (ECOSUR-257) with four pairs of lateral teeth, and one pair of basal spines larger than the rest of the spines present in medial spike ( Fig. 10L–M View FIGURE 10 ).
Median ridge long, with marginal eyespots. Median organ rhomboid, pale, with a light brown line extending from the median ridge to the central region of the median organ (ECOSUR-P3207; Fig. 9F View FIGURE 9 ).
Some specimens with fragments of the tube, with white sand (ECOSUR-255; Fig. 10N View FIGURE 10 ).
Etymology. This species is named after Dr. Sergio I. Salazar-Vallejo, a polychaete specialist, who has shared with me his knowledge and has given me his support. The species name is a noun in the genitive case ( ICZN 1999, Art. 31.1.2).
Distribution. Yucatán Peninsula and Florida coast, in intertidal to sublittoral (5 m) waters. On rocks, sand flat, or associated with Millepora hydrozoans.
Remarks. Sabellaria salazari n. sp. belongs to the group of species having penicillate medial spike and resembles Sabellaria sp. A sensu Uebelacker, 1988 from the Gulf of Mexico and S. wilsoni Lana & Gruet, 1989 from Brazil.
Sabellaria salazari n. sp. has outer paleae with a longer medial spike, ½ of the total length of the outer palea. Uebelacker (1988) did not describe the length of the medial spike of Sabellaria sp. A , but illustrated the outer paleae with a spike almost longer than the blade of outer paleae (Uebelacker 1988: Fig. 49-6c). Lana & Gruet (1989) described S. wilsoni with a short spike, ¼–1/6 as long as outer paleae, and reexamined Uebelacker’s material describing a longer medial spike, 1/3-1/5 as long as outer paleae of Sabellaria sp. A .
According to Lana & Gruet (1989), Sabellaria sp. A has 5–6 pairs of lateral spines on outer paleae, and S. wilsoni has 3–6. Sabellaria salazari n. sp. has 3–5 pairs of lateral spines; however, it is very similar to Sabellaria sp. A because it has wider lateral spines on one side, whereas S. wilsoni has lateral spines of similar size on both sides. Sabellaria sp. A could belong to S. salazari n. sp. but it is necessary to review Uebelacker’s material to confirm this.
Although Lana & Gruet (1989) described S. wilsoni as having “middle paleae directed upward, nearly straight and just slightly bent inwards…” it is necessary to consider the angle between the blade and the handle to estimate the inclination of the paleae. Lana & Gruet (1989) illustrated the middle paleae with a convex angle of 140° to 150° between the blade and handle, greater than that of S. salazari n. sp. (125° to 140°); however, the position of the paleae of S. wilsoni is ventral and not lateral.
Liñero-Arana (2013) recorded S. wilsoni for Venezuela and included scanning electron microscope photographs of paleae. However, the middle palea has a smaller angle (~100° in lateral view) than my estimate for S. wilsoni (140–150° in ventral view). In addition, the inner paleae are wider in the Venezuelan specimens (2–3 times longer than wide) than that of S. wilsoni (5 times longer than wide). Therefore, it is recommendable to review the type material of S. wilsoni to clarify the morphology of middle paleae and to consider the angle between the blade and the handle in the opercular paleae for future descriptions.

| R |
Departamento de Geologia, Universidad de Chile |
| UF |
Florida Museum of Natural History- Zoology, Paleontology and Paleobotany |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |