
Sabellaria floridensis Hartman, 1944
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5048.2.3 |
|
publication LSID |
lsid:zoobank.org:pub:CBD94592-5647-4F52-A8CF-4990DE8574D8 |
|
DOI |
https://doi.org/10.5281/zenodo.5555039 |
|
persistent identifier |
https://treatment.plazi.org/id/03D0AE23-516C-FFD8-FF73-FE01FEC81705 |
|
treatment provided by |
Plazi |
|
scientific name |
Sabellaria floridensis Hartman, 1944 |
| status |
|
Sabellaria floridensis Hartman, 1944 View in CoL
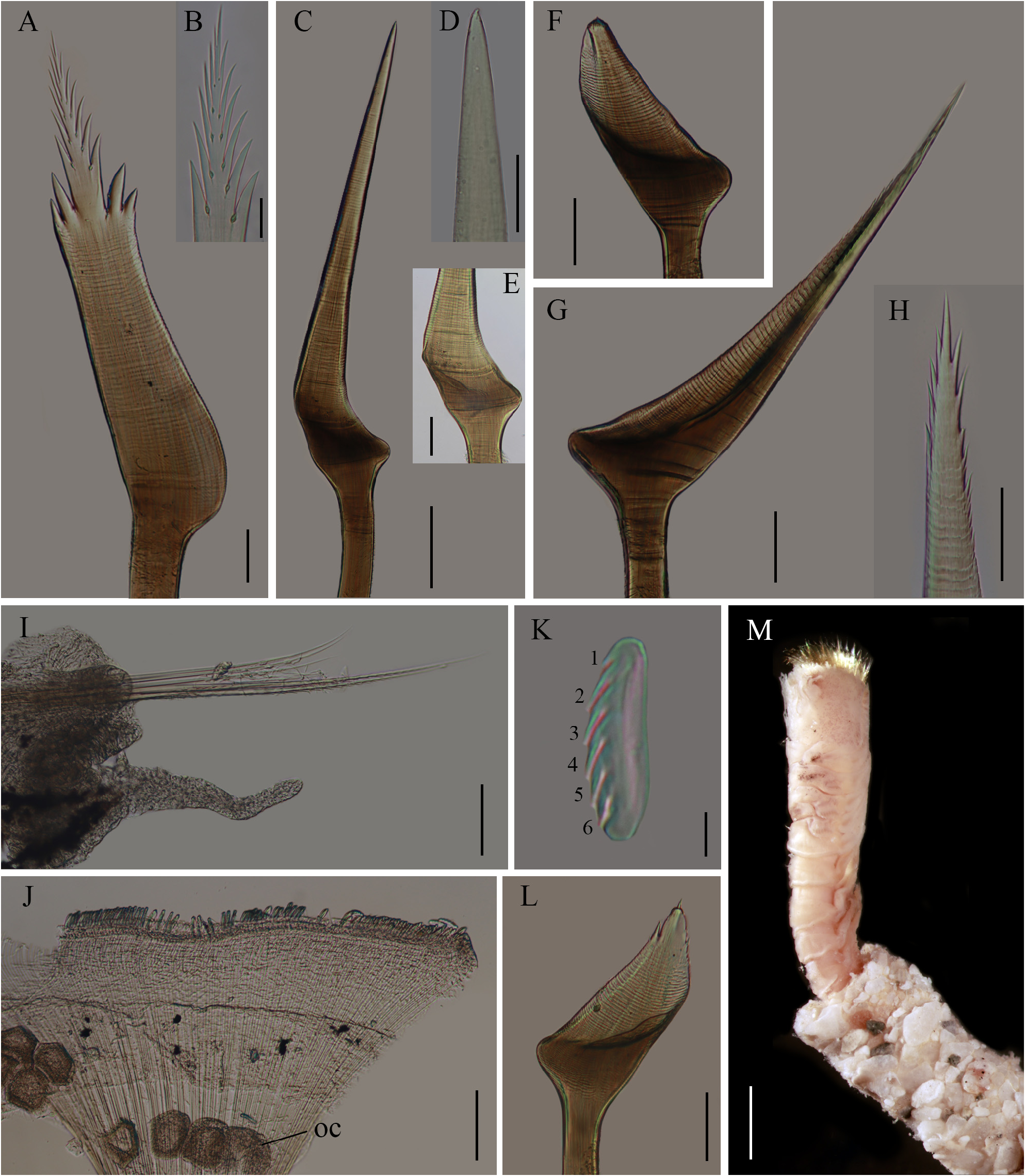
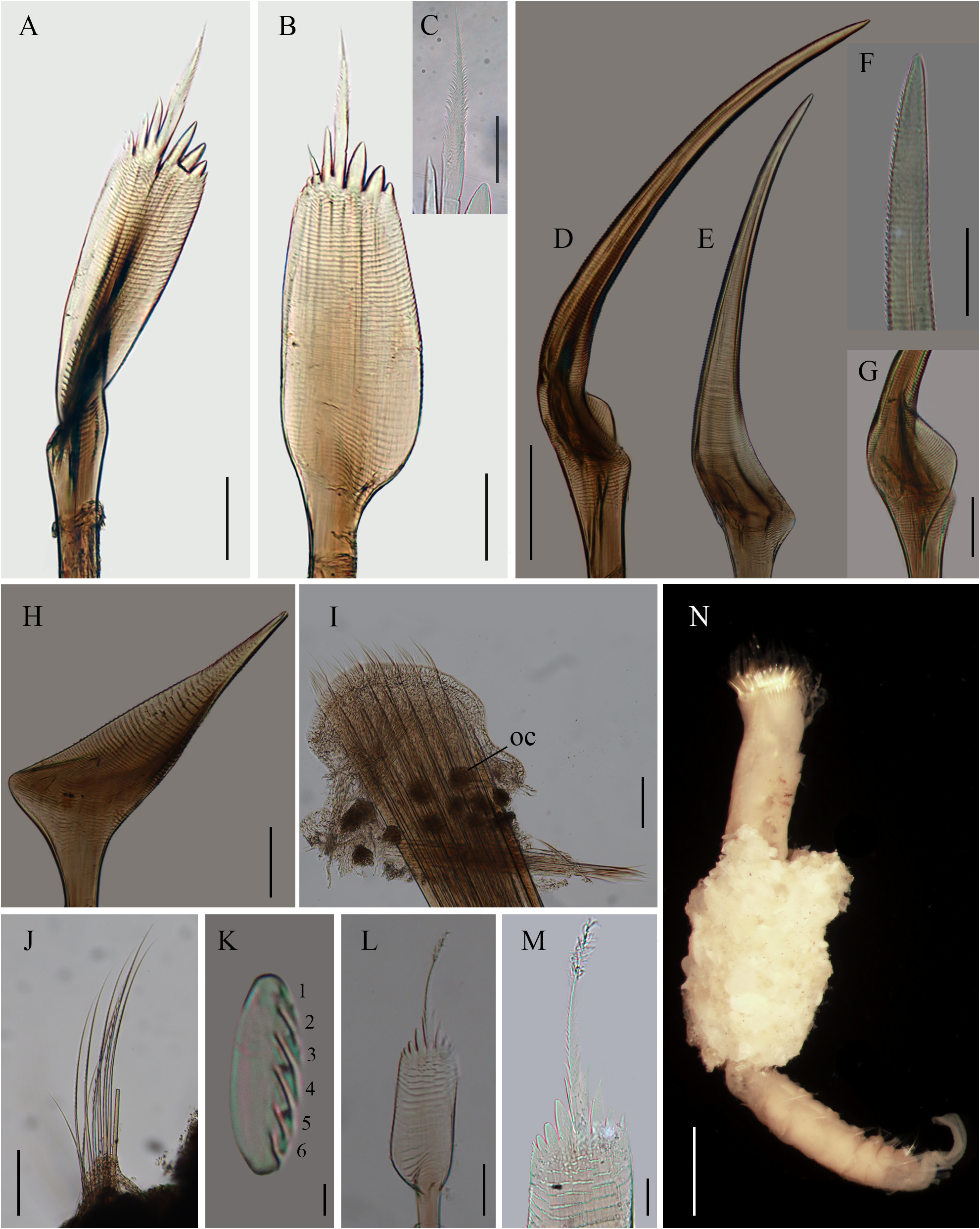
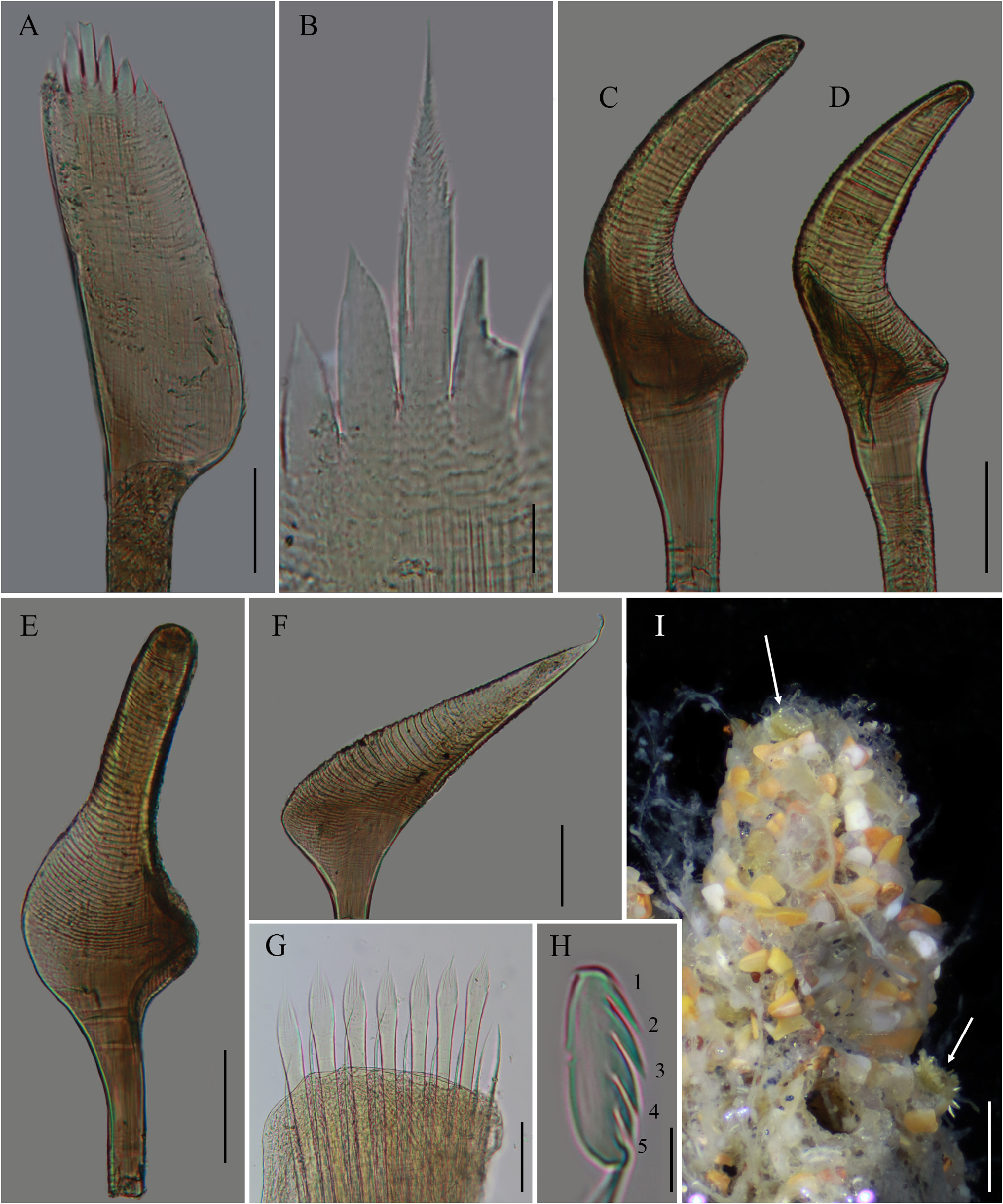
Figures 6A–F View FIGURE 6 ; 7A–M View FIGURE 7 ; 8A–E View FIGURE 8
Sabellaria floridensis Hartman, 1944: 345–346 View in CoL , pl. 31, figs. 37–41. Type locality: Lemon Bay , Florida.
Sabellaria floridensis View in CoL .— Rioja 1946: 196, figs. 2–9 (Tecolutla Beach, Veracruz, in mollusk shells, associated with S. vulgaris beaufortensis View in CoL ); Uebelacker 1984: 49.8, fig. 49–4 (Gulf of Mexico); Kirtley 1994: 58–60, figs. 4.10.1–4.10.3 (North Carolina to Florida, Gulf of Mexico, associated with P. caudata View in CoL ).
Material examined: 39 specimens. Campeche: ECOSUR-P3197 , one spec. ( Champotón , on rock, 4–5 m, February 16, 1999, coll. J. R. Bastida-Zavala & S.I. Salazar-Vallejo). ECOSUR-P3198 , two spec. ( Champotón , February 16, 1999, coll. J. R. Bastida-Zavala & S.I. Salazar-Vallejo). ECOSUR-P3199 , one spec. ( Sabancuy , July 28, 1984, coll. S.I. Salazar-Vallejo). ECOSUR-P3200 , eight spec. ( Champotón , February 16, 1999, coll. J. R. Bastida-Zavala & S.I. Salazar-Vallejo). ECOSUR-P3201 , five spec. ( Champotón , 3 m, February 15, 1999, coll. J. R. Bastida-Zavala & S.I. Salazar-Vallejo). ECOSUR-P3202 , 12 spec. ( Champotón , 6–6.5 m, February 16, 1999, coll. L. Cosgalla Delgado & V. Reyes Galindo). ECOSUR-P3203 , one spec. ( Champotón , February 14, 1999, coll. J. R. Bastida-Zavala & S.I. Salazar-Vallejo). ECOSUR-P3204 , one spec. (Campamento Tortugero Punta Xen, Champotón , May 24, 2005, on rock, 20 m from the coast, 1 m, coll. S.I. Salazar-Vallejo & L.F. Carrera-Parra). Florida: UF283 , six spec. (Seahorse Key, Cedar Keys , 29°06’N, 83°04’W, March 31, 1973) GoogleMaps .
Description. Complete specimen (ECOSUR-P3199), broken in the last segments, mature female. Body colorless, 15 mm long, 2 mm wide, with three parathoracic segments, 21 abdominal segments, caudal peduncle 3 mm long ( Fig. 6A–D View FIGURE 6 ).
Opercular crown and opercular stalk partially fused ( Fig. 6C View FIGURE 6 ). Opercular disc oblique, three rows of amber paleae; outer row with 24–25 paleae per side, middle row with 6–7 long and 7–8 short paleae per side, inner row with 15–16 paleae per side. Outer paleae with two lateral teeth; blade flat, oblong, basally wider, almost five times longer than wide; thecae transverse with finely serrate margins. Blade bent towards the outside of the opercular crown; blade and handle at an angle of almost 170° ( Fig. 7A View FIGURE 7 ). Medial spike straight, ½ as long as blade, with six pairs of lateral teeth bearing a wedge-shaped process at the base ( Fig. 7B View FIGURE 7 ). Middle row with long and short interspersed paleae. Long paleae geniculate, slightly longer than outer paleae; blade almost straight at an angle of 155°, thecae transverse with finely serrate margins ( Fig. 7C View FIGURE 7 ); tips blunt, smooth ( Fig. 7D View FIGURE 7 ). Blade base concave with a short lateral expansion ( Fig. 7E View FIGURE 7 ). Short middle paleae geniculate, with concave blade directed outwards; thecae transverse with finely serrate margin; tips blunt, smooth ( Fig. 7F View FIGURE 7 ). Blade basally with a thin lateral expansion. Inner paleae geniculate, ½ as long as middle dorsal paleae ( Fig. 7G View FIGURE 7 ); concave blade directed inwards, distally serrate; thecae transverse with serrate margin ( Fig. 7H View FIGURE 7 ).
Opercular peduncle colorless ( Fig. 6A, C View FIGURE 6 ). Two nuchal spines on left lobe and three spines on right lobe ( Fig. 6C, E View FIGURE 6 ). Fifteen pairs of opercular papillae. Tentacles and palps pale ( Fig. 6B View FIGURE 6 ). Median ridge as long as the opercular stalk, with marginal dark eyespots. Median organ globular. Building organ pale.
Thorax without notochaetae. Chaetiger 1 with a pair of neuropodia with capillary chaetae. Chaetiger 2 with intensely dark pigment laterally, capillary neurochaetae, and a pair of branchiae ( Fig. 6B View FIGURE 6 ).
Parathorax with three segments, all with one pair of branchiae. First parathoracic segment darkly pigmented laterally. Second parathoracic segment paler. Third segment colorless. Notopodia with 10 lanceolate chaetae, 11 capillary chaetae. Neurochaetae lanceolate, thinner than notochaetae.
Abdominal segments colorless, branchiae decreasing in size posteriorly; absent in the last five chaetigers. First two segments with long neuropodial cirri ( Fig. 10I View FIGURE 10 ). Neurochaetae verticillate. Notopodia with a series of uncini with six transverse rows of teeth ( Fig. 7J–K View FIGURE 7 ). Caudal peduncle colorless, cylindrical, slightly annulate ( Fig. 6D View FIGURE 6 ).
Parathoracic and abdominal parapodia with oocytes, each about 60 µm in diameter ( Fig. 7J View FIGURE 7 ).
Variation. Body 3–21 mm in total length, 0.4–2.5 mm wide, 10–24 abdominal segments, caudal peduncle 0.5–3 mm long. The number of opercular paleae varied after the size of specimens: 16–25 outer paleae per lobe, 4–8 middle long paleae per lobe, 4–8 middle short paleae per lobe, and 10–15 inner paleae per lobe. Outer paleae with 2–3 lateral teeth. Medial spike with 4–7 pairs of lateral teeth.
Only one specimen (ECOSUR-P3197) had short middle paleae with serrate tips, and terminal spine ( Fig. 7L View FIGURE 7 ). The marginal spines decrease in size from dorsal to ventral paleae.
Some specimens with fragments of tubes made of sand and shell fragments (ECOSUR-P3198; Fig. 7M View FIGURE 7 ).
Distribution. North Carolina to Campeche ( Uebelacker 1984) on rocks, intertidal to 6.5 m depth.
Remarks. Sabellaria floridensis Hartman, 1944 and S. bella Grube, 1870 are two closely related species that share outer paleae with denticulate medial spike, and middle paleae with long and short forms.
Grube (1870) made a brief description of S. bella from Desterro (now Florianópolis), Southern Brazil. Augener (1934) re-described the type material and illustrated the opercular paleae. Augener mentioned that outer paleae have 6–8 teeth with a central tooth larger than the others. The central tooth (or medial spike) has a fine, hair-like tip, with about 10 pairs of lateral teeth. The inner paleae have lateral teeth in the distal third of the blade.
Hartman (1944) described S. floridensis from Lemon Bay, Florida with “22 pairs of prolonged outer paleae with two larger serrations on a side and a minute tooth at the outer base; six pairs of alternating long and short middle paleae; and 11 pairs of inner paleae terminating distally in a serrated edge”. Also, Hartman (1944) recorded S. bella from Beaufort, North Carolina, and Independencia Bay, Peru, and she indicated in her identification key ( Hartman 1944: p. 339–340), that the differences between these species were in the tip of the inner paleae, being entire in S. bella and serrate in S. floridensis . However, Augener (1934) described S. bella with serrate inner paleae as well. Thus, S. bella and S. floridensis share outer paleae with lateral teeth and denticulate medial spike, middle paleae with long and short forms, and inner paleae with serrate tip.
Gruet & Lana (1988) commented on the differences in the opercular paleae of S. bella and S. bellis Hansen, 1882 , both species from southern Brazil. Here, I focus on the description of S. bella . According Gruet & Lana (1988), Sabellaria bella has 20 pairs of outer paleae with 2–3 lateral spines (or teeth) on each side of a central or medial spike; 13 pairs of middle paleae, and 12 pairs of inner paleae.
Some differences can be seen between the redescriptions of S. bella by Augener (1934) and by Gruet & Lana (1988). The medial spike has 5–6 lateral teeth in Gruet & Lana (1988) and not 10 pairs as Augener (1934) described. The long middle paleae have a more robust base, three times wider than the median blade region as seen in Augener’s illustrations (1934: Fig. 31c), and almost twice as wide in the photographs and illustrations by Gruet & Lana (1988: Figs. 1F View FIGURE 1 , 2D View FIGURE 2 ). On the other hand, the short middle paleae illustrated by Augener (1934: Fig. 31b) are more concave, with recurved margins and with an abrupt sharp point at the apex, whereas Gruet & Lana (1988: Figs. 1D–E View FIGURE 1 , 2C View FIGURE 2 ) show slightly concave paleae, with straighter margins and sharp apex. The inner palea has a serrate tip; however, these inner paleae have a bent blade in base, an angle of around 130° between handle and blade in Gruet & Lana (1988: Figs. 1H View FIGURE 1 , 2E View FIGURE 2 ), while Augener (1934: Fig. 31d) does not mention anything about the bending, but the angle between handle and blade is larger than 137°. The differences observed can be due to the position where the paleae illustrated were taken from, lateral in Augener’s schemes, and ventral, upper, or another position as in Gruet & Lana’s photographs and drawings.
Both Augener (1934) and Gruet & Lana (1988) examined one or two specimens, so it is difficult to infer the morphological variations that may be present in the opercular paleae of S. bella . Other records of S. bella have also included few specimens in their descriptions ( Kirtley 1994: two spec.; Santos et al. 2011: four spec.; Liñero-Arana 2013: five spec.), and it has even been mentioned that S. bella is a solitary species ( Santos et al. 2011). However, it is recommended that S. bella should be revised to expand its description and include variations in the opercular paleae to better delimit the specific diagnostic characters.
Although Kirtley (1944) reviewed the type material of Sabellaria bella and S. floridensis , he neither emphasized the differences in the two species nor provided illustrations for S. bella , but took those of Gruet & Lana (1988) to illustrate the opercular morphology. Besides, there are some contradictions between his identification key to species of Sabellaria and the corresponding species descriptions. For example, Kirtley described S. bella with denticulate medial plume on the outer paleae, although in the key this species is grouped with those lacking this feature. Therefore, it is not possible to deduce which features he considered diagnostic for S. bella and S. floridensis .
Regarding S. floridensis, Kirtley (1994) mentioned that there are morphological variations in the middle paleae, and that rigorous analysis is needed. He stated that some short middle paleae were slightly bent; however, in none of the specimens reviewed herein was this pattern identified. This is probably due to the number of samples reviewed (33 lots), the size of the specimens, or the localities of collection, as Kirtley (1994) included specimens from North Carolina to the Gulf of Mexico, Venezuela, and Panama (Atlantic coast), whereas most of the specimens reviewed here are from the Gulf of Mexico.
Based on the above, some morphological differences between S. bella and S. floridensis ( Figs. 7–12 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 View FIGURE 11 View FIGURE 12 ) can be determined. The medial plume of the outer paleae is slightly longer in S. floridensis (blade 1.6 times longer than medial spike, Figs. 7A View FIGURE 7 ; 12A View FIGURE 12 ) than in S. bella (blade twice longer than median spike, Fig. 8F View FIGURE 8 ). The short middle paleae have blunt tips in S. floridensis ( Figs. 7F View FIGURE 7 ; 12C View FIGURE 12 ) and sharp tips in S. bella ( Fig. 8H View FIGURE 8 ). The inner paleae are seen bent from the base, with marginal spines 1/3–¼ as long as distal middle spine in S. bella ( Gruet & Lana 1988: p. 33, Figs. 1H–I View FIGURE 1 ; 8J View FIGURE 8 ) and straight paleae with marginal spines half as long as the distal middle spine in S. floridensis ( Figs. 7H View FIGURE 7 ; 12E View FIGURE 12 ).
Sabellaria floridensis has been found from the same aggregation as S. salazari n. sp. Both species are distinguished mainly by the characteristics of the medial spike in the outer paleae and the morphology of the middle paleae. Sabellaria salazari n. sp. has penicillate medial spike ( Fig. 10A–C View FIGURE 10 ) and only middle paleae in long-form ( Fig. 10D–E View FIGURE 10 ), while S. floridensis has denticulate medial spike ( Fig. 7A–B View FIGURE 7 ) and middle paleae in long and short forms ( Fig. 7C–F View FIGURE 7 ).




| R |
Departamento de Geologia, Universidad de Chile |
| V |
Royal British Columbia Museum - Herbarium |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Sabellaria floridensis Hartman, 1944
| Chávez-López, Yessica 2021 |
Sabellaria floridensis
| Hartman, O. 1944: 346 |