
Achaeta macroampullacea, Dózsa-Farkas & Felföldi & Nagy & Hong, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4496.1.27 |
|
publication LSID |
lsid:zoobank.org:pub:7C536E1E-5D5A-4E2D-9E4F-28F3CEA9664C |
|
DOI |
https://doi.org/10.5281/zenodo.5950201 |
|
persistent identifier |
https://treatment.plazi.org/id/03D3D43A-E44F-FFA6-2580-F91FFE65FF1B |
|
treatment provided by |
Plazi |
|
scientific name |
Achaeta macroampullacea |
| status |
sp. nov. |
Achaeta macroampullacea View in CoL sp.n.
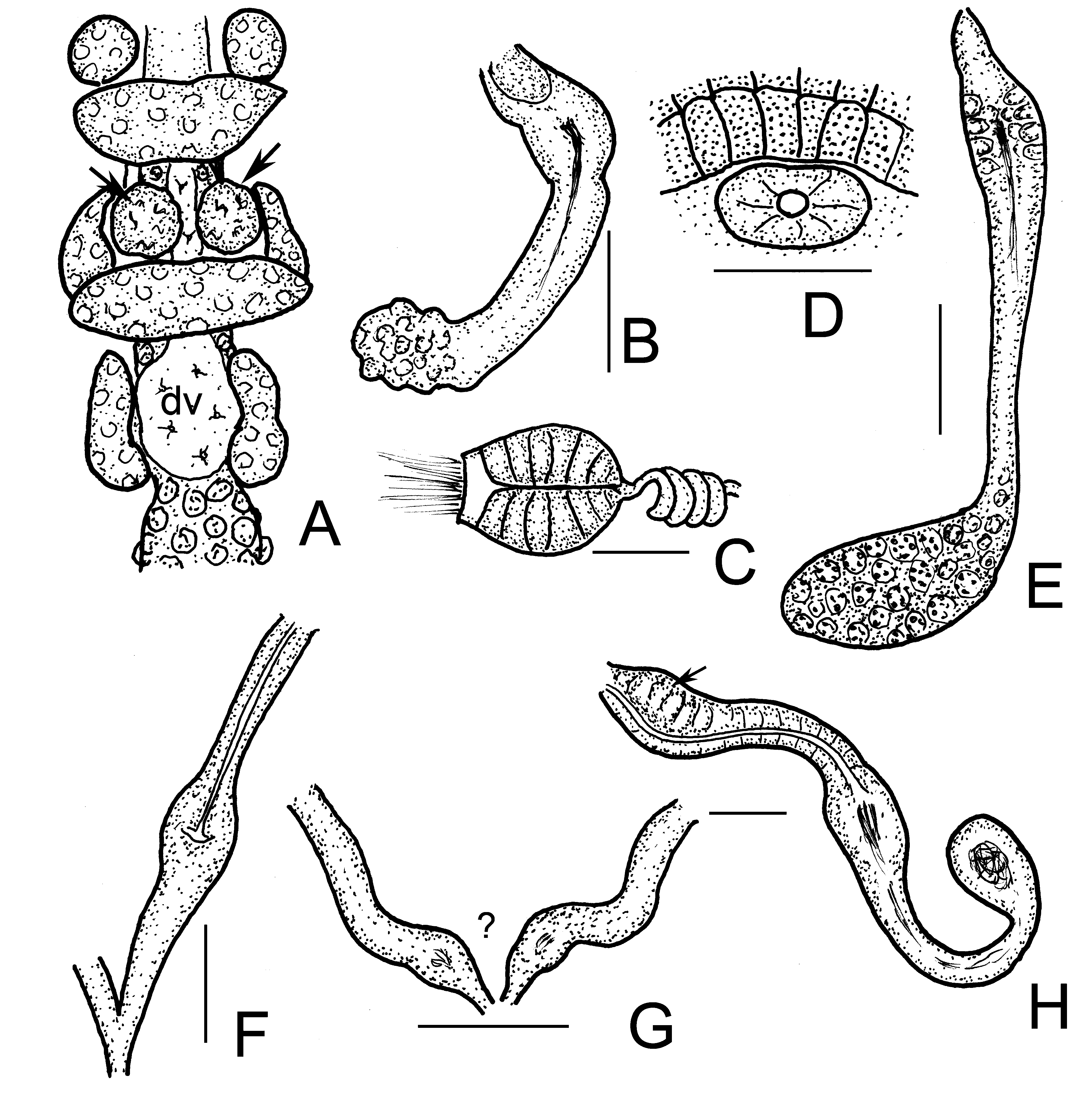
( Figures 3 View FIGURE 3 , 4 View FIGURE 4 , 5D–E View FIGURE 5 )
Type material. Holotype: NIBRIV0000810586, slide No. 2353, adult, stained whole mounted specimen. Type locality: site 12, Mt. Hallasan , Jeju Island , Korea, soil and litter layers of mixed forests, near to Gwaneumsa temple, N 33˚25'41.5", E 126˚33'47.2", 551 m asl, 29.09.2016, leg. Y. Hong. Paratypes (in total 14 stained, adult specimens on slides and 12 specimens in 70% ethanol): NIBRIV0000810587, slide No. 2320, NIBRIV0000811383, slide No. 2337, Soil in Yongnuni-orums, Gujwa-eup, Jeju Island, Korea, 26.10.2016, leg. Y. Hong, P . 114.1.1–114.1.4, slide No. 2275, 2354–2355 a, b, site 11, P.114.2.1–114.2.7, soil in Yongnuni-orums, Gujwa-eup, Jeju Island, Korea, 26.10.2016, leg. Y. Hong, P . 114.3, slide No. 2356, soil in bottom of Yongnuni- orums, Gujwa-eup , Jeju Island , Korea, 26.10.2016, leg. Y. Hong. In 70% ethanol: P.114.4 soil in Yongnuni-orums , Gujwa-eup, Jeju Island, Korea, 26.10.2016, leg. Y. Hong, 12 specimens .
Further material examined. 8 specimens investigated in vivo, 2 of them processed for DNA analysis, one of them (spcm No. 1137, see Table 3): bottom of Yongnuni-orums, Gujwa-eup , Jeju Island, N 33°27’30.94”, E 126°49’54.90”, 193 m asl, 26.10.2016, leg. Y. Hong. GoogleMaps
Etymology. Named after the large spermathecal ental reservoir of ampulla.
Diagnosis. The new species can be recognized by the following combination of characters: (1) small worms (2.5–4 mm long and 190–220 µm wide at clitellum in vivo), segments 20–26; (2) no pyriform glands; (3) clitellum developed only laterally: in dorso-lateral area the hyalocytes more or less in two longitudinal rows with granulocytes bordered, ventro-laterally the granulocytes ordered in about three longitudinal rows; (4) dorsal blood vessel from VII; (5) all pharyngeal glands connected dorsally with ventral lobes, the first ventral lobes in IV the largest, secondary ventral glands absent; (6) two pairs of preclitellar nephridia in 6/7–7/8; (7) pars tumida of midgut from XIV–XX, extending over 2–4 segments; (8) sperm funnel small, barrel-shaped, collar narrower than funnel body; (9) male pores in XII, ventro-laterally, no glandular body, only the pore surrounded by small inconspicuous glands; (10) spermathecae free, extending into VI–VII, ental reservoir large.
Description. Small worm. Holotype 3.7 mm long, 200 µm wide at VIII and 230 µm at clitellum (fixed), 28 segments. Length of paratypes 2.5–4.0 mm, width 150–200 µm at VIII and 19 0–220 µm at clitellum in vivo, length of fixed specimens 2.3–3.8 mm, width 150–230 µm at VIII and 150–260 µm at clitellum; segments 20–30. No pyriform glands. One pair of small epidermal glands dorsally. Body wall dorsally 12–17 µm thick, cuticle 5–7 µm, ventrally 10–12 µm and 2.5–5 µm, respectively, fixed ( Fig. 3E View FIGURE 3 ). Clitellum in XII–1 /2 XIII, developed only laterally, dorso-lateral areas with hyalocytes in 2 disordered longitudinal rows ( Figs. 3G–H View FIGURE 3 ) between granulocytes. The dorsal interruption about 40–50 µm wide in vivo, dorsal borderline consisting of granular cells only in one longitudinal row, latero-ventrally the granulocytes in three or four longitudinal rows ( Fig. 3H View FIGURE 3 ). Hyalocytes 17–20 µm high and 10–14 µm wide, granulocytes 18–22 by 8–14 µm, smaller at dorsal borderlines and between the rows of hyaline cells (16 by 10 µm) in vivo. The ventral interruption about 40–60 µm wide. Head pore on top of prostomium. Spermathecal pores at 4/ 5 in lateral position. Male pores in XII.
Brain egg-shaped 80–100 µm long and about 2 times longer than wide ( Fig. 3A,B View FIGURE 3 ). Prostomial ganglion small about 15–23 µm long, fixed ( Fig. 3B View FIGURE 3 ). All pharyngeal glands at 4/5–6/7 united dorsally, all with ventral lobes. The first ventral lobes conspicuous ( Fig. 3C,D View FIGURE 3 ), secondary ventral lobes absent. Septa in 4/5–6/7 thickened. Two pairs of preclitellar nephridia at 6/7, 7/8 ( Fig. 3F View FIGURE 3 ), constricted by septum; length ratio anteseptale: postseptale 1: 2, postseptale tapers gradually into efferent duct, with terminal vesicle. Dorsal blood vessel from VII, blood colourless. Coelomocytes about 30–48 µm long in vivo, flat, cytoplasm light brown, spindle shaped in side view ( Figs. 4A,B View FIGURE 4 ). Oesophageal appendages spongy in V, well developed without canal in IV. Chloragocytes yellowishbrown, about 14–23 µm long in vivo. Midgut pars tumida XIV–XX (occupying 2–4 segments) ( Figs. 3I –J View FIGURE 3 ). Sperm funnel small, barrel-shaped, 70–120 µm long in vivo (50–87 µm, fixed), about 1.5–2 times longer than wide, collar narrower than funnel body and 11–15 µm high ( Figs. 4C–E View FIGURE 4 ). Sperm duct 7–9 µm thick in vivo (5–7 µm, fixed) mostly wound into a regular spiral ( Fig. 4C View FIGURE 4 ). Spermatozoa 48–75 µm, heads 22–30 µm long in vivo (45–55 µm and 10–20 µm, fixed). Seminal vesicle absent. Male copulatory organs small, separate ventro-laterally, no glandular body or bursa, only the pore surrounded by small inconspicuous glands (41–54 µm long, 31–38 µm wide in vivo, 35–45 µm long and 20–33 µm wide, fixed) ( Figs. 4F View FIGURE 4 , 5D View FIGURE 5 ). Spermathecae ( Figs. 4G View FIGURE 4 , 5E View FIGURE 5 ) free. The ectal duct (20–30 µm long, 12–20 µm wide) widens out to a glandular dilation of ampulla (30–60 µm long, 20–30 µm wide). The contractile narrow connecting tube (60–120 µm long, 10–13 µm wide) expands into a large ental reservoir reaching segment VI or VII ( Figs. 4G–H View FIGURE 4 ). Sometimes one in VI the other in VII. This ental reservoir is 50–130 µm long and 30–55 µm wide in vivo. In the reservoir sperm absent, full of globules, probably oil drops (which is supported by the fact that in fixed worms this part is barely visible, since alcohol presumably dissolves oil drops).
Distribution and habitat. In Korea, Mt. Hallasan, few specimens at site 11 and 12, but more common in Yongnuni-orums, Gujwa-eup, Jeju Island in mixed forests.
Differential diagnosis. Among the previously described 6 Achaeta species without pyriform glands and spermathecae reaching segments VI–VII, A. littoralis Lasserre, 1968 and A. iridescens Christoffersen, 1979 are much larger (35–40 segments, length 15–20 mm and 55–60 segments with 13.5–14.5 mm, respectively, vs. 20–26 segments and 2.5–4.0 mm in A. macroampullacea sp. n.). A. hallensis has two secondary ventral lobes of the pharyngeal glands and a large seminal vesicle (vs. absent in the new species). In A. macrocyta Christensen & Dózsa-Farkas, 1999 the origin of the dorsal vessel is from VIII and in A neotropica from XIII–XV (vs. from VII). The main distinguishing feature of A. hanagarthi is only one pair of preclitellar nephridia in 7/8 (vs. two pairs in 6/ 7 and 7/ 8 in the new species).


| DNA |
Department of Natural Resources, Environment, The Arts and Sport |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |