
Eunice tovarae, Carrera-Parra, Luis F. & Salazar-Vallejo, Sergio I., 2011
|
publication ID |
https://doi.org/ 10.5281/zenodo.208034 |
|
DOI |
https://doi.org/10.5281/zenodo.6189014 |
|
persistent identifier |
https://treatment.plazi.org/id/03E087AA-FD06-053A-FF74-FC47629CFCB8 |
|
treatment provided by |
Plazi |
|
scientific name |
Eunice tovarae |
| status |
sp. nov. |
Eunice tovarae View in CoL n. sp.
Figure 3 View FIGURE 3
Eunice filamentosa Fauchald 1970:31 View in CoL –33, Pl. 3, Figs. c–g (syn., non Grube & Örsted in Grube, 1856).
Material examined. Type material: Holotype ECOSUR-104, Pacific, Mexico: Sinaloa, Dock of R/V El Puma, 23° 10ʹ 53.4ʺ N, 106° 25ʹ 27.56ʺ W, Mazatlán, 20 Oct 2010, fouling over concrete dock pilings, 0.5 m. Paratypes: ECOSUR-105, Pacific, Mexico: Sinaloa, Dock of R/V El Puma, 23° 10ʹ 53.4ʺ N, 106° 25ʹ 27.56ʺ W, Mazatlán, 14 Aug 2008, fouling over concrete dock pilings, 0.5 m. ECOSUR-106 (ECOSUR-OH-P0103), Pacific, Mexico: Sinaloa, cruise ship port, 23°11ʹ43ʺ N 106°24ʹ50.9ʺ W, Mazatlán, 8 Mar 2008, col. María Ana Tovar-Hernández ( MATH), fouling over concrete dock pilings, 0.5 m .. ECOSUR-107 (ECOSUR-OH-P0111) Pacific, Mexico: Sinaloa, cruise ship port, 23°11ʹ43ʺ N 106°24ʹ50.9ʺ W, Mazatlán, 8 Mar 2008, coll. MATH, fouling over concrete dock pilings, 0.5 m. ECOSUR-108 (ECOSUR-OH-P0112) Pacific, Mexico: Sinaloa, cruise ships port, 23°11ʹ43ʺ N 106°24ʹ50.9ʺ W, Mazatlán, 8 Mar 2008, coll. MATH, fouling over concrete dock pilings, 0.5 m. Additional material: Pacific, Mexico: Sinaloa, ECOSUR (4) same data as holotype. ECOSUR (2), Dock of R/V El Puma, 23° 10ʹ 53.4ʺ N, 106° 25ʹ 27.56ʺ W, Mazatlán, 14 Aug 2008, fouling over concrete dock pilings, 0.5 m. ECOSUR-OH- P0091 (1), ferry dock, 23° 11ʹ 10.9ʺ N, 106° 25ʹ 12.2ʺ W, Mazatlán, 11 Mar 2008, coll. MATH, fouling over metallic dock pilings, 0.7 m.
Description. Holotype (ECOSUR-104) complete with 251 chaetigers, L10 = 5.8 mm, W10 = 2.4 mm, TL = 8.5 cm. Anterior region of body with convex dorsum and flat ventrum; body depressed from chaetiger 14. Prostomium shorter than peristomium, prostomial lobes frontally rounded, dorsally inflated; median sulcus deep ( Fig. 3 View FIGURE 3 A). Prostomial appendages in an evident horseshoe pattern, median antenna isolated by gap. Palps thicker than antennae, digitiform, reaching middle of first peristomial ring. Antennae tapering; lateral antennae reaching middle of first chaetiger; median antenna reaching anterior margin of second chaetiger. Palpophores and ceratophores wider than long, but the former wider than the latter. Palpostyles and ceratostyles without articulation. Eyes subspherical, lateral between palp and lateral antenna. Peristomium with first ring five times longer than second ring, separation between rings dorsal and ventral; peristomial cirri tapering, exceeding middle of first peristomial ring; without articulation ( Fig. 3 View FIGURE 3 A).
Maxillary apparatus with five pairs and one single maxillae; MF = 1+1, 4+4, 6+0, 2+7, 1+1, –+–. MIII part of distal arc, blunt teeth, distal tooth slightly shorter. Left MIV with both teeth on distal end followed by a long edentate plate ( Fig. 3 View FIGURE 3 F). Right MIV with blunt teeth of similar size. MVI reduced to an edentate plate.
Branchiae from chaetiger 22 to last chaetiger, always longer and thicker than dorsal cirri; one branchial filament in chaetigers 22–32, then pectinate ( Fig. 3 View FIGURE 3 D), increasing gradually up to six filaments in the last third of body. Prechaetal lobes inconspicuous, transverse folds in chaetigers 1–70, in posterior chaetigers following outline of chaetal lobes. Postchaetal lobes rounded, slightly longer than chaetal lobes in chaetigers 1–19, shorter in chaetigers 20–56, inconspicuous from chaetiger 57. Chaetal lobes elongated, as long as wide, with aciculae emerging dorsal to midline; in first 17 chaetigers with a papilla covering acicular tip ( Figs. 3 View FIGURE 3 B–E).
Notopodial cirri without articulation, tapering; first three slightly thicker, becoming thinner gradually towards posterior parapodia; all of similar length. Ventral cirri in chaetigers 1–3 digitiform, thick; in chaetigers 4–74 with swollen base as transverse welt with short digitiform tip; digitiform from chaetiger 75, increasing in length to be longest in the most posterior chaetigers, always shorter than notopodial cirri ( Figs. 3 View FIGURE 3 B–E).
Chaetae limbate supracicular; pectinate heterodonts in all chaetigers, with up to 14 teeth. Compound falcigers bidentate, in anterior chaetigers with long, thin blade, proximal tooth thicker, directed laterally; distal tooth very thin, sharp, directed upward ( Fig. 3 View FIGURE 3 G). Falcigers in posterior chaetigers with short, wide blade, proximal tooth much thicker, directed laterally; distal tooth very short, directed laterally ( Fig. 3 View FIGURE 3 H). Aciculae black, in chaetigers 1– 5 with tips expanded to one side ( Fig. 3 View FIGURE 3 J), laterally expanded in chaetigers 6–237, T-shaped ( Fig. 3 View FIGURE 3 K), last 13 chaetigers with aciculae tapering, with small mucro ( Fig. 3 View FIGURE 3 L); always one per chaetiger. Subacicular hooks from chaetiger 17, bidentate, black, tip slightly translucent; always one per chaetiger, except for replacements. Proximal tooth much larger than distal tooth; strongly curved, more evident in posterior chaetigers ( Fig. 3 View FIGURE 3 M); distal tooth directed laterally.
Pygidium with two pairs of anal cirri without articulation; dorsal pair as long as the last four chaetigers, ventral pair very short.
Variation. Material examined varies in L10 from 2.9 to 6.0 mm, and in W10 from 1.4 to 2.6 mm and varies in the following features: Subacicular hooks begin from chaetiger 16 to 19, branchiae start from chaetiger 20 to 26, while the last chaetiger with ventral cirri with swollen base varies from 61 to 79. Although there were few specimens, these features seem to be size-dependent. There is a correlation between the start of subacicular hooks and branchiae, the former always start before branchiae. The maximum number of filaments varies from three to five, the best development observed in the last third of body (the higher filaments number is present in specimens with L10 greater than 5.0 mm).
Barcode. Nucleotide sequences between 618–663 bp of the section of COI gene used for barcoding were obtained from three paratypes (ECOSUR-OH-P0103, ECOSUR-OH-P0111, ECOSUR-OH-P0112). The average evolutionary divergence over the three sequence pairs was of 0.1%.
Type locality. Mazatlán, Sinaloa (Mexican Pacific coast).
Etymology. This species is being named after Dr. María Ana Tovar-Hernández, in recognition of her many publications on sabellid polychaetes and her sampling efforts on harbor fauna, which included the specimens used for this description.
Distribution. Mexican Pacific coast. This species has been referred to as E. filamentosa and has been found from the northern part of the Gulf of California to Oaxaca, along the Mexican Pacific coast. Some other records under the same name from Panama or Galápagos might belong to this newly described species.
Discussion. Among Eunice species, E. tovarae n. sp. resembles E. filamentosa and E. denticulata in several features. They all have a right MIV with two teeth in the distal end, followed by a long edentate plate, a late start of branchiae, ventral cirri with swollen bases as a transverse welt with short digitiform tips, bidentate subacicular hooks, and aciculae distally hammer-shaped in at least some parapodia.
However, there are several differences among these species; E. tovarae n. sp. and E. filamentosa differ from E. denticulata in the shape of both MIII and MIV on the right side (MIVr). The former species have MIII with blunt teeth, distal tooth slightly larger and MIVr with blunt teeth of similar size, while E. denticulata has MIII and MIVr with sharp curved teeth, with medial teeth longer. Furthermore, these two species differ from E. denticulata by having the region with the greatest branchial development in the last third of the body, and branchiae are always larger that notopodial cirri; whereas in E. denticulata the best branchial development is in the middle of body and the branchiae are shorter than notopodial cirri in most posterior parapodia.
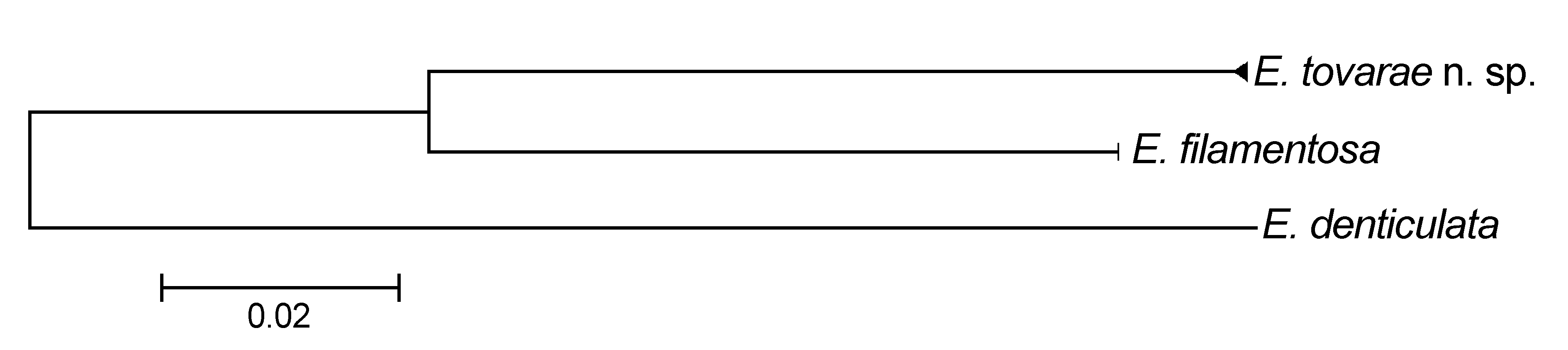
On the other hand, E. tovarae n. sp. differs from E. filamentosa by having black aciculae and subacicular hooks instead of being amber, as in E. filamentosa . An additional difference refers to features of the chaetal lobes: in E. tovarae n. sp. they are rounded, elongated, and shorter than postchaetal lobe in most anterior chaetigers; whereas in E. filamentosa they are auricular, and always larger than the postchaetal lobe. Another difference is in the length of the body region with VCSB; in E. tovarae n. sp. this region ends about chaetigers 61–78, and in E. filamentosa it ends about chaetigers 33–66. Although this feature is size-dependent, there are significant differences between both species if we compare specimens of similar size; thus, specimens of E. tovarae n. sp. with L10= 4.0– 5.2 mm have VCSB until chaetiger 65–78; while specimens of E. filamentosa with a L10= 4.1–5.0 mm have VCSB until chaetiger 40–59. Furthermore, E. tovarae n. sp. has composite falcigers with blades twice as long in anterior chaetigers than in posterior ones; whereas E. filamentosa has composite falcigers with blade of similar length in all chaetigers. The morphological differentiation of the three species was supported by the barcoding data. Eunice tovarae n. sp. has a 20.7% genetic divergence from E. denticulata and a 12.9% with respect to E. filamentosa ( Fig. 4 View FIGURE 4 ). The main variation in the sequences of these species was in the GC% of the third codon position. In E. tovarae n. sp. it varied from 35.8 to 36.5%, in E filamentosa it varied from 33.2 to 33.6%, while for E. denticulata it was 45%. Although the objective of this work is not to establish a phylogenetic relationship of Eunice species, it seems that E. filamentosa is more closely allied to E. tovarae n. sp. than to E. denticulata . Hebert et al. (2003) considered that sequence divergences between species average 6.8% for congeneric taxa, and are higher for more distantly related taxa. Furthermore, Jones et al. (2008) and Vrijenhoek et al. (2009) found sequence divergences among closely related Osedax polychaete species from 8.4%.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.