
Rhynchelmis (Rhynchelmoides) bolinensis, Fend, Steven V. & Lenat, David R., 2010
|
publication ID |
https://doi.org/ 10.5281/zenodo.196882 |
|
DOI |
https://doi.org/10.5281/zenodo.6206479 |
|
persistent identifier |
https://treatment.plazi.org/id/03EEF043-2A35-6B60-A1D3-FED3FB69A02E |
|
treatment provided by |
Plazi |
|
scientific name |
Rhynchelmis (Rhynchelmoides) bolinensis |
| status |
sp. nov. |
Rhynchelmis (Rhynchelmoides) bolinensis View in CoL n. sp.
( Figures 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 )
Holotype: USNM 1142469, a longitudinally dissected worm, stained in hematoxylin and slide mounted.
Type locality: USA, North Carolina, Chatham Co. , small seep beside Pokeberry Creek, 7 Jan 2009, collected by D.R. Lenat.
Paratypes: The type locality, 7 Jan 2009: USNM 1142470-1142472, 3 dissected. 18 Dec 2008: USNM 1142473-1142474, 1 sagittally sectioned, 1 transversely sectioned.
Other material: The type locality, 28 Feb 2007: 4 dissected, several in alcohol; all partially or postmature. 18 Mar 2007: 1 whole mount, several in alcohol, none mature. 18 Dec 2008: 10 dissected, 1 sagittally sectioned, several in alcohol. 7 Jan 2009: 10 dissected, several in alcohol. 23 Jan 2008: 2 dissected, 1 sagittally sectioned, 1 transversely sectioned. Halifax Co.: Beech Swamp at SR 1100, 30 Dec 2008: 3 dissected, 2 mature and several immature in alcohol. 28 Jan 2010: 3 dissected, slightly post-mature. Orange Co.: seep on unnamed tributary to Bolin Creek, 22 Jan 2009: 2 dissected. Bolin Creek and Dry Gulch (a tributary of Bolin Creek), 0 6 March 2009: 2 dissected, several immature in alcohol. Unnamed tributary to Little River at Star, 18 Mar 2007: several immature. All collected by D.R. Lenat.
Etymology. Named for Bolin Creek, in recognition of conservation efforts by the Town of Carrboro in protecting this stream.
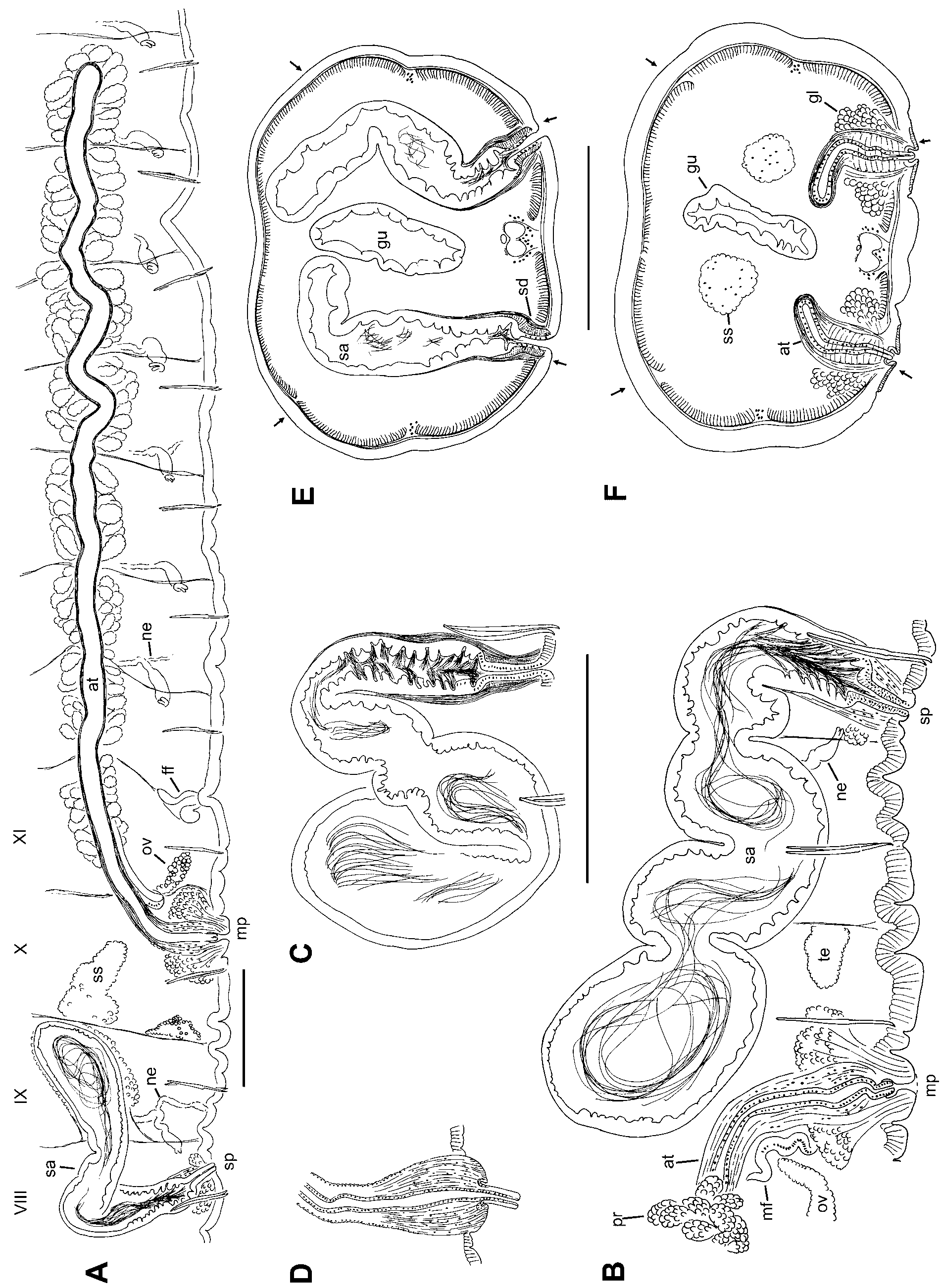
Description. Length of preserved worms 38–48 mm; segments 114–135; diameter 1.3 (0.8–1.6) mm in X, maximum diameter 1.4 (0.9–1.8) mm. Prostomium with a distinct filiform proboscis, about 1 mm long at the type locality, but up to 2.5 mm in some Beech Swamp worms ( Fig. 1 View FIGURE 1 A–C). Secondary segmentation usually a narrow anterior ring in about IV–VIII, variable and indistinct behind clitellum. Epidermis about 15–20 μm thick in preclitellar area, up to 40–70 μm in clitellum, and 10–15 μm posteriorly. Longitudinal muscle 20–30 μm thick, usually thicker ventrally than dorsally; circular muscle about 5 μm. Clitellum mid-VIII or IX to at least XV, but usually weakly developed to XVII; thickest in X–XIII. Chaetae simple-pointed, moderately sigmoid, nodulus approximately 1/3 the distance from tip (0.3–0.42); the inner chaeta in each pair slightly longer than the outer ( Fig. 1 View FIGURE 1 D). In clitellar region, length of ventral chaetae 256 (180–355) μm, dorsals slightly shorter, 213 (170–288); in posterior segments, 150–260 μm, with dorsals about equal. Pharynx from II–V, without a distinct dorsal pad; columnar cells about equally developed dorsally and ventrally. Pharyngeal glands usually IV–VI. Brain in the peristomium.
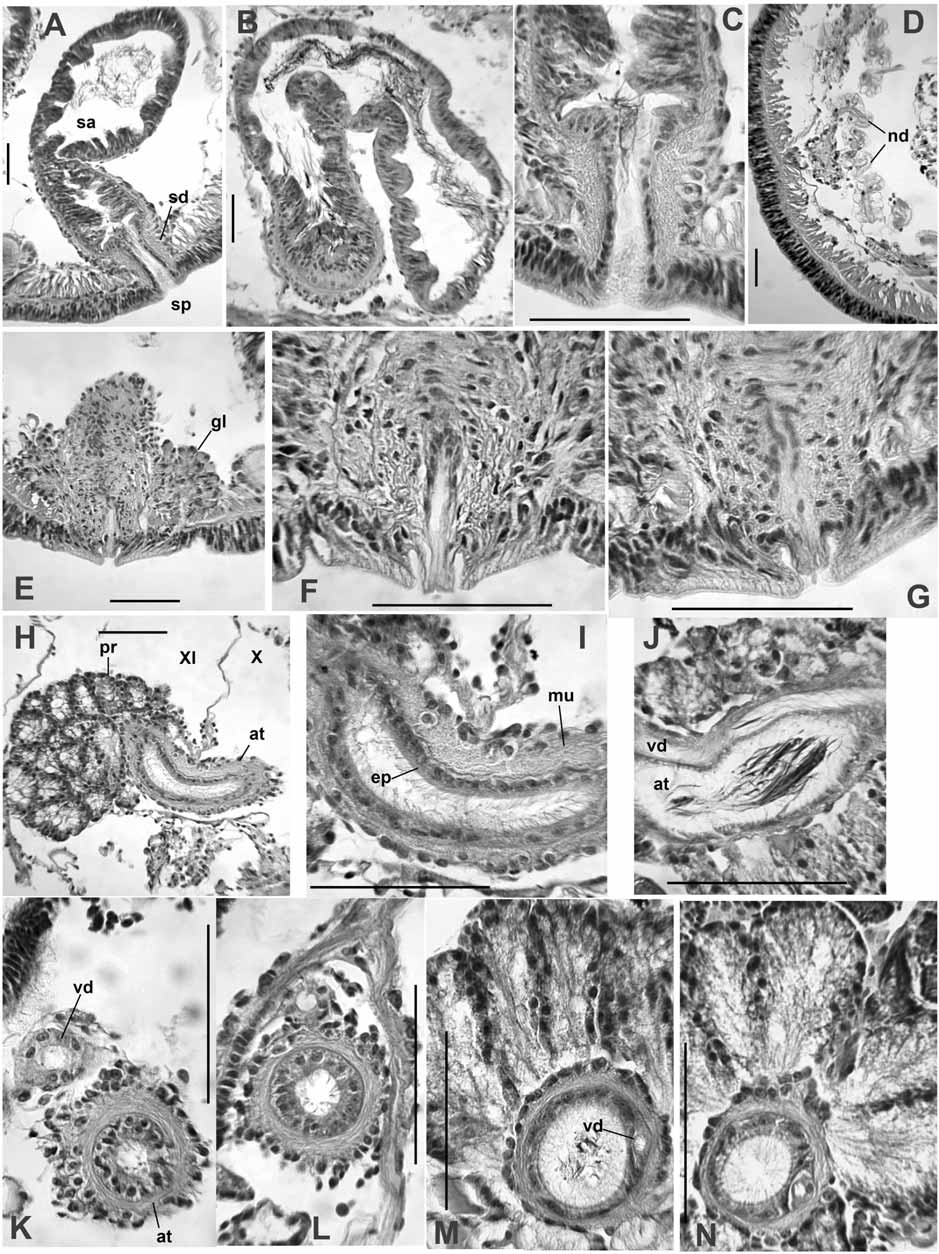
First nephridia usually paired on 7/8 and/or 8/9; next pair usually on 12/13; nephridia usually paired in segments posterior to XIII, but sometimes absent in middle and posterior segments. Each nephridium with small anteseptal funnel (length about 25–40 μm), followed by a largely postseptal, ovate expansion (length about 85–120 μm, diameter 35–70 μm), and a long, convoluted duct which forms a narrow loop; the conjoined ducts extend dorsally along the anterior lateral blood vessel ( Fig. 3 View FIGURE 3 D), then ventrally along a posterior lateral vessel, and may continue posteriorly, ventral to the gut. The ectal end of the duct terminates in an inconspicuous nephropore anterior to the ventral chaetae in the originating segment, without forming a distinct vesicle. The expanded portion may be entirely postseptal, or part may extend through the septum; in this case, the funnel is usually twisted.
Two anterior forks of ventral blood vessel join near 6/7. One pair of lateral blood vessels joins the dorsal vessel in the posterior part of each segment, beginning in II. These vessels are commissures, joining the ventral vessel in segments anterior to about XIII (or its forks, anterior to VII); they are long and sinuous anterior to IX; shorter, with 5 or more long, blind branches in segments posterior to X; in IX and X they may loop posteriorly through many segments within sperm and egg sacs. A second pair of lateral blood vessels, having long, blind branches in anterior part of VIII and posterior segments; up to 5–10 branches per lateral vessel. One branch of each anterior lateral vessel joins perivisceral sinus at ventral side of gut posterior to about XV; in segments posterior to about L, the posterior lateral vessels also have a branch to the gut. Posterior to about LX or LXX, branches of lateral vessels shorter and more numerous, often with bulbous tips. Perivisceral sinus and chloragogen well-developed on gut beginning in about VII; posterior lateral vessels covered with chloragogen near dorsal vessel in anterior segments, entirely covered in posterior segments; all anterior lateral vessels covered with chloragogen. Blood glands at perivisceral sinus, at junction of small blood vessels from ventral vessel; up to 3 per segment from about IX to at least XV.
Testes paired in X, relatively small, not extending to mid-segment. Ovaries in XI, not extending beyond mid-segment ( Fig. 2 View FIGURE 2 A). Sperm sacs extend posteriorly to XXII–XLIII. Egg sacs to XXV–XLV. Female pores intersegmental on 11/12, in line with ventral chaetae; female funnels simple, cup-shaped, 130–180 μm high. Spermathecae paired in VIII; atria paired in X; both male and spermathecal pores posterior to, and in line with the ventral chaetae ( Fig. 2 View FIGURE 2 A–B, E–F). Male pores on low, oval mounds 120–190 μm wide, slightly wider than long.
Spermathecae are elongate-sacciform, typically folded within VIII, but in many cases extending into IX or X, within septal sacs on 8/9 and 9/10 ( Fig. 2 View FIGURE 2 A–B); vestigial sacs present even when spermathecae do not penetrate septa ( Fig. 2 View FIGURE 2 A). Elongate ampulla in two parts ( Fig. 2 View FIGURE 2 B–C). The ental portion irregular, 1250 (850– 2100) μm long, by about 200–400 μm wide; epithelium irregular, usually 15–30 μm thick, without obvious sorptive vacuoles; muscle layer indistinct; lumen with sparse, unordered sperm ( Fig. 3 View FIGURE 3 B). The shorter ectal portion, about 200–300 μm long by 100–180 μm wide, has a thicker (35–50 μm), deeply folded or incised epithelium, usually with sperm heads lined up in folds, and a distinct, outer muscle layer to 5 μm thick ( Fig. 2 View FIGURE 2 B, C; 3A, C). Ectally, the spermatheca terminates in a compact duct, 85–130 μm long; duct is a narrow tube of thin epithelium surrounded by a thick (12–15 μm) layer of circular-transverse muscle fibers ( Fig. 3 View FIGURE 3 C). Spermathecal pores without obvious accessory glands.
Male funnels on 10/11 only, 150–230 μm high, usually not strongly convoluted, but may extend posteriorly a short distance within the sperm sac. Vasa deferentia enter X directly, without forming a loop in the postatrial segment or penetrating septum 10/11. Vas deferens usually joins the atrium within the sperm sacs in the anterior part of XI, near the beginning of the prostate layer; vas runs under the atrial muscle coat to join the atrial lumen at the ental end ( Fig. 3 View FIGURE 3 K–N). Free portion of vas deferens very short, less than 400 μm long, width 22–36 μm.
Atria elongate, tubular, extending posteriorly within the sperm sacs to XV–XIX ( Fig. 2 View FIGURE 2 A). The short (500 to 800 μm long) ectal portion of each atrium extends at most to mid-XI; this section has a 10–14 μm thick layer of circular muscle, cuboidal epithelium 7–12 μm thick, and lacks prostates; diameter 65–90 μm, lumen narrow (15–25 μm) ( Fig. 3 View FIGURE 3 H, I, K, L). The much longer (2000–3900 μm) ental portion usually begins in XI; this part of the atrium is unevenly tubular, diameter 67–112 μm, not expanded entally. Histology of the ental portion similar for its entire length: an uneven muscle layer about 6–10 μm thick; a variable epithelial layer about 3–10 μm; and a wider lumen (about 40–55 μm), usually filled with sperm ( Fig. 3 View FIGURE 3 H, J, M, N). Prostate cells granular, in pyriform clusters 100–170 μm long; these glands are very densely packed over the entire ental part of the atrium ( Fig. 3 View FIGURE 3 M, N).
Ectal end of atrial duct surrounded by a thick layer of mostly circular muscle fibers interspersed with longitudinal fibers; resultant penial structure about 220–300 μm long by 120–150 μm wide. The duct (about 40 μm wide) may be slightly convoluted within the muscular layer, or extended to form a short terminal penis; duct epithelium is cuboidal. Penes are short extensions (to 50 μm) of the atrial duct, and may protrude slightly or be enclosed within a pad of non-clitellar tissue forming the ectal mound ( Fig. 3 View FIGURE 3 F–G); extended penis on a post-mature worm was 70 μm long ( Fig. 2 View FIGURE 2 D). Male pore internally surrounded by a loose cluster of petiolate gland cells, either single or in small bundles resembling prostates; the narrow ends of these gland cells insert on the ectal pad. Glands extend to about 200 μm high; the entire cluster about 200–400 μm wide ( Fig. 3 View FIGURE 3 E).
Remarks. Rhynchelmis bolinensis is similar to R. alaskana , but it lacks the prominent midventral glands in anterior segments of that species. More subtle differences include the histology of the spermathecal ampulla, the more highly developed penial structures, the more tubular atrium, and the presence of nephridia in VIII and/or IX. Rhynchelmis alaskana appears restricted to northern Alaska, but another related species lacking a spermatheca-gut connection, R. saxosa , occurs from Montana to Washington and Oregon. In addition to the abovementioned characters, R. bolinensis differs from R. saxosa in having extensive, branched, lateral blood vessels.
The mid-December collections included mated and unmated worms; most specimens collected in early January had sperm in the spermathecae, and the spermathecal ampullae were much larger than those collected in December. By late January, most specimens appeared to be resorbing their reproductive organs, and by early March only a few worms had vestigial reproductive pores.
Rhynchelmis bolinensis was collected from two very different types of habitat: small streams and seeps in parts of the Carolina Slate Belt (Chatham/Orange counties) and a coastal plain swamp in Halifax County. These areas, do, however, have some characteristics in common: poorly drained clay soils, accumulations of leaves in winter months, and a tendency to dry up during summer months. Both areas became completely dry prior to our 2008/2009 collections, suggesting this species may require a dry period in order to complete its life cycle. Large numbers of these worms were found in the headwaters of Bolin Creek and in Dry Gulch (a tributary of Bolin Creek), 0 6 March 2009; in this habitat the species was associated with leaf packs rather than the stream sediment. Rhynchelmis bolinensis was not present in several earlier collections at the swamp site, suggesting a very patchy distribution. Additional immature worms collected at other sites are provisionally assigned to this species, based on the relatively long proboscis and nephridia in VIII and IX.

| USNM |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.