
Onuphis farensis, Gil, João & Machado, Margarida, 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3860.4.3 |
|
publication LSID |
lsid:zoobank.org:pub:872A1117-F6C6-4471-A5B9-9034FD8D3A99 |
|
DOI |
https://doi.org/10.5281/zenodo.5689578 |
|
persistent identifier |
https://treatment.plazi.org/id/03FB422E-6C3C-A666-FF03-730052711839 |
|
treatment provided by |
Plazi |
|
scientific name |
Onuphis farensis |
| status |
sp. nov. |
Onuphis farensis View in CoL sp. nov.
(figs. 2–6)
Onuphis (Nothria) geophiliformis View in CoL (not Nothria geophiliformis Moore, 1903 View in CoL )— Ibáñez 1972: 26 –28, fig. 2 (Sandbank of Cabezuela, Bay of Cádiz, Spain); Ibáñez 1973a: 128 ( Isla Cristina and Cádiz, Spain); Ibáñez 1973b: 43 –45, gráfico 12, cuadro 13 ( Isla Cristina and Cádiz, Spain).
Nothria geophiliformis View in CoL (not Nothria geophiliformis Moore, 1903 View in CoL )— Campoy 1982: 557 (previous records: Sandbank of Cabezuela, Cádiz, and Isla Cristina, Spain).
? Nothria geophiliformis View in CoL (not Nothria geophiliformis Moore, 1903 View in CoL )— Dexter 1992: 79 (Ria de Alvor).
? Onuphys geophyliformis (not Nothria geophiliformis Moore, 1903 View in CoL )— Mucha & Costa 1999: 371 (Ria de Aveiro and/or Sado Estuary, Portugal).
Onuphis View in CoL sp. nov. — Gil 2011: 544 –545 (Ria Formosa, Portugal).
Material examined. PORTUGAL, Ramalhete (Faro, Ria Formosa), 37º00’20.9”N, 07º58’28.1”W, WGS84 datum, intertidal sand. Areia Intertidal 4, 27 /03/2001 (G.3761): HOLOTYPE MB29-000332, 1 c adult, broken in two pieces, with Ciliophora, with fragment of tube. Areia Intertidal, 25/11/1999 (G. 162): PARATYPE MB29-000333, 1 af adult. Areia Intertidal, 07/02/2000 (G. 162): PARATYPE MB29-000334, 1 c adult with a membranous tube with attached small sand grains of different sizes, posterior end damaged. Areia Intertidal 3, 27 /03/2001 (G. 3754): PARATYPE NMW.Z.2014.010.0001, 1 af adult with Ciliophora, 2 mf; PARATYPE NMW.Z.2014.010.0002, 2 af juveniles. Areia Intertidal 6, 27 /03/2001 (G. 3770): PARATYPE NMW.Z.2014.010.0003, 1 af adult, with Ciliophora. Areia Intertidal 9, 03 /07/2001 (G. 3736): PARATYPE MB29-000335, 2 af juveniles, with Ciliophora. Areia Intertidal 1, 28 /01/2002 (G. 3810): PARATYPE SMF 23896, 1 af adult, with Ciliophora. Areia Intertidal 3, 28 /01/2002 (G. 3821): PARATYPE MB29-000336, 1 af adult, 1 mf, with Ciliophora. Areia Intertidal 6, 28 /01/ 2002: PARATYPE MB29-000337, 1 af adult, 1 mf, with Ciliophora. Areia Intertidal, 12/06/2003: PARATYPE MB29-000338, 2 af adults, 20 af juveniles and young adults of different sizes, some with Ciliophora, one with tube.
Description. Based on the holotype, complemented with data from paratypes.
Complete adult (holotype MB29-000332) 47.5 mm long, for 163 chaetigers, broken in two pieces. Live worms not observed, body flesh colored in alcohol, without visible colour patterns. Body cylindrical up to approximately chaetiger 4, being dorsoventrally flattened from chaetiger 5 rearwards (posterior to the pharynx) and with segments shorter in length, tapering towards pygidium within the last 20 chaetigers (fig. 2A–B).
Prostomium small, subtriangular with a rounded anterior margin. A pair of oval to obclavate frontal lips. Palpophores and ceratophores arranged in semicircle. Palps reaching chaetigers 1–2, with 6–9 rings on palpophores. Lateral antennae reaching chaetiger 4–6, with 9–11 rings on each ceratophore, being always longer than median antenna. Median antenna reaching chaetiger 3–4, with 5–7 rings on ceratophore. Nuchal organs not observed. Eyes not visible, apparently absent. Peristomium slightly shorter than first chaetiger, bearing two slender tentacular cirri at the outer anterior margin (right one missing on holotype), generally about half the length of the peristomium.
Jaw apparatus description based on a single specimen (paratype MB29-000333), consisting of dorsal maxillae and ventral mandibles. Maxillae (fig. 3A) weakly sclerotized, translucent with some darkened margins. Carriers triangular, about as long as MI. Maxillary formula as MI: 1+1, MII: 9+9, MIII: 10+0, MIV: 7+10, MV: 1+1. Proximal tooth very small in both maxillae MII, and in MIII. Mandibles (fig. 3B) slightly shorter than maxillae, with slender and poorly sclerotized shafts, and slightly calcified cutting plate, with irregular edge. Lateral shelves not observed. Cutting plate with the two heavily sclerotized protomandibles completely embedded in the translucent surrounding mandibles.
First 4 chaetigers slightly displaced ventrally, with longer dorsal and ventral cirri, and postchaetal lobes (fig. 2A–B, fig. 4A). First 2 chaetigers about as long as wide, with parapodia directed anteriorly. Proceeding chaetigers becoming quickly wider and shorter (fig. 2A). Dorsal cirri present throughout the body, being very long on anterior chaetigers, but diminishing gradually there after and becoming short for the largest part of the body, up to the last chaetigers (fig. 4A–D). Ventral cirri cirriform in the first 4 chaetigers (fig. 4A), being replaced by glandular pads thereafter. Occasionally, chaetiger 5 showing a transitory ventral cirrus, consisting of a slightly longer pad (figs. 2B, 4B). Digitiform postchaetal lobes elongate and conical, fully developed in the first 8 chaetigers (fig. 4A–B), then reducing progressively (fig. 4C), becoming small and papilliform by chaetiger 18, being present up to chaetiger 23–33 in the biggest adults (fig. 6B), and absent posteriorly (fig. 4D). From about chaetiger 8–10 the postchaetal lobe displaces ventrally, becoming subchaetal, with limbate chaetae emerging from the superior base of the lobe (fig. 4C). Interramal papillae absent.
Simple strap-like branchiae first present from chaetiger 5 (figs. 2A, 4B), appearing at the inner base of the dorsal cirri, being intially shorter or only marginally longer than the corresponding dorsal cirri (fig. 4B), but quickly surpassing it, reaching the dorsal midline of the body (fig. 4C–D). Branchiae absent in the last 50–60 chaetigers.
Chaetigers 1–4 with bi– and tridentate pseudocompound hooks with blunt tips, numbering 4 (rarely 3, 5 or 6) per parapodium (fig. 5A–C). Majority tridentate, sometimes with a tiny and difficult to see basal denticle, but bidentate hooks not uncommon (although not present in every specimen). Some tridentate hooks show a bifid middle (fig. 5D) and proximal denticles (fig. 5E, S3), or an extra tiny denticle between the distal and middle denticles (fig. 5E, S1). Additional completely formed hooks are present inside the body, at the base of the parapodia. Simple limbate chaetae with narrow wings from chaetiger 5, absent in anterior chaetigers bearing pseudocompound hooks. Pectinate chaetae flat and oblique with approximately 10 teeth (fig. 5G), normally present from chaetiger 5 (can be difficult to detect, but always appearing no later than chaetiger 6 or 7 in adult specimens), numbering one or two per parapodium. Very small capillary chaetae present in most, if not all, chaetigers, including the first four, which posses pseudocompound hooks. A pair of bidentate subacicular hooks present from chaetiger 9, each with a short, squared hood, first emerging from the base of the anterior region of the postchaetal lobe (fig. 4C–D, fig. 5F).
Pygidium bearing 2 pairs of ventrally inserted anal cirri, upper pair longer than lower pair, dorsal rim crenulated (fig. 2C).
Tube membranous, encrusted with poorly-sorted sand grains.
Remarks on juveniles. Some adult characters are invariable in juveniles, indicating they are fixed in an early stage of ontogeny. Branchiae always appear on chaetiger 5, and subacicular hooks are normally present from chaetiger 9, except in 3 juvenile specimens, in which only one hook is present in one of the parapodia of chaetiger 8, but being present in both parapodia from chaetiger 9. Very young juveniles do not show pectinate chaetae, but when they do they are normally present in the first branchial chaetigers, usually at chaetiger 5, as in the adults (fig. 6A). Bi – and tridentate pseudocompound hooks are also present, but with a higher proportion of bidentate hooks in relation to adults.
However, other important characters show a bigger variability between juveniles and adults, and seem to be related with size or with the ontogeny of the species. These are: the number of ventral cirri, the number of chaetigers with pseudocompound hooks, and the number of chaetigers with postchaetal lobes. Younger juveniles bear ventral cirri only in the first 3 chaetigers (instead of 4), and pseudocompound hooks may also be present only in the first 3 chaetigers. Alternatively chaetiger 4 may possess a mixture of hooks and limbate chaetae. In general, specimens with pseudocompound hooks present only in the first 3 chaetigers, lack ventral cirri entirely or are very reduced on the 4th chaetiger, indicating the link between these two characters, at least in some onuphids, as previously noticed by Fauchald (1982b).
Completely developed postchaetal lobes vary in number, ranging between 3–8 and absent on chaetiger 9 in younger juveniles, or present only as decreasingly sized bulges up to chaetiger 11–21 in older juveniles and young adults.
Methyl green staining pattern. Based on holotype MB29-000332, and paratypes MB29-000336 and MB29- 0 0 0 337.
Palpophores, palps and ceratophores, and frontal, upper and lower lips stained deeply, unlike the ceratostyles which remained unstained. Peristomium stained near the anterior margin, peristomial cirri strongly stained. Anterior region of body with parapodia, dorsal and ventral cirri, postchaetal lobes and branchial filaments strongly stained. Posterior region with branchial filaments and dorsal cirri unstained.
Ventral glandular pads stained deeply, with posterior half darker than anterior. From the middle region rearwards, anterior and posterior bases of each parapodium with strongly stained dark blue dots, forming vertical stripes anterior and posterior to the glandular pads. Those posterior to the pads being longer and darker.
Dorsum in the anterior region with broad transverse pale stripes, formed by two thinner lines, uniting both parapodia of the same segment, and becoming paler rearwards.
Ventrum of first chaetiger with a V-shaped stripe formed by thinner lines uniting both parapodia, leaving a paler area in the anterior region of the segment, just behind the lower lip. On the second chaetiger the stripe is more anteriorly placed, and from the third segment it becomes central in the segment. From about chaetiger 10 it turns into 2 separate bands: an incomplete anterior one at the level of the glandular pads, interrupted at the midline and a complete posterior one, uniting the region just posterior to the glandular pads. This pattern fades posteriorly. In the middle and posterior region of the body a broken longitudinal midventral line appears.
Posterior region and pygidium without strong staining patterns, just some discontinuous thin dorsal transversal lines in the last chaetigers, formed by dark dots.
Etymology. The specific epithet ‘ farensis ’ refers to the type locality of the species, being near the town of Faro, in Ramalhete, located in the coastal lagoon and Natural Park of Ria Formosa (Southern Portugal).
Type locality. Southwestern Iberian Peninsula: Ramalhete, near the town of Faro ( Portugal, 37º00’20.9”N, 07º58’28.1”W, WGS84 datum), at the Ria Formosa mesotidal coastal lagoon (Ria Formosa Natural Park), in an intertidal sandbank.
Ecology. Intertidal sandbanks of very fine to coarse sand, and in meadows of Zostera noltii Hornemann , in coastal lagoons of the Gulf of Cádiz ( Portugal and Spain).
Adults were collected in Ramalhete (Faro, Portugal) between 1999 and 2003 throughout the whole year, whilst juveniles were only collected in the first half of the year (2nd May 2000, 27th March 2001, 3rd July 2001, and 12th June 2003) during the same period. Densities of the species in Ramalhete oscillated between 8.8 ind/m2 (27th November 2000) and 97.3 ind/m2 (12th June 2003).
In Cabezuela (Cádiz, Spain) the species was recorded in the intertidal zone, in a Zostera noltii Hornemann meadow (as Zostera nana Roth ) ( Ibáñez 1972).
Ibáñez (1973b) references the species from five intertidal stations from Cabezuela (Cádiz, Spain) and Isla Cristina (Huelva, Spain), without giving more specific details about the locality of each station. One of the stations is located in a Zostera noltii Hornemann meadow (as Zostera nana Roth ), and is probably the same station mentioned in Ibáñez (1972) from Cabezuela (Cádiz). The density of the species was stated to be 182 ind/m2 at a station within well sorted very fine sand. However, figures of lower than 25 ind/m2 at the other four stations were noted, within moderately to well sorted very fine sand (3 stations), and in moderately sorted coarse sand (1 station), in salinities ranging from 33.993‰ to 35.796‰.
Distribution. Atlantic Ocean, Gulf of Cádiz, Southwestern Iberian Peninsula: Ramalhete, Ria Formosa (Faro, Portugal; type locality), Sandbank of Cabezuela (Cádiz, SW Spain; Ibañez 1972, 1973b), and Isla Cristina (Huelva, SW Spain; Ibañez 1973a, 1973b). Other earlier records from Portugal may also refer to O. farensis sp. nov., but need to be confirmed: Ria de Alvor ( Dexter 1992; as Nothria geophiliformis ), and Ria de Aveiro and/or Sado Estuary ( Mucha & Costa 1999; as Onuphys geophyliformis ).
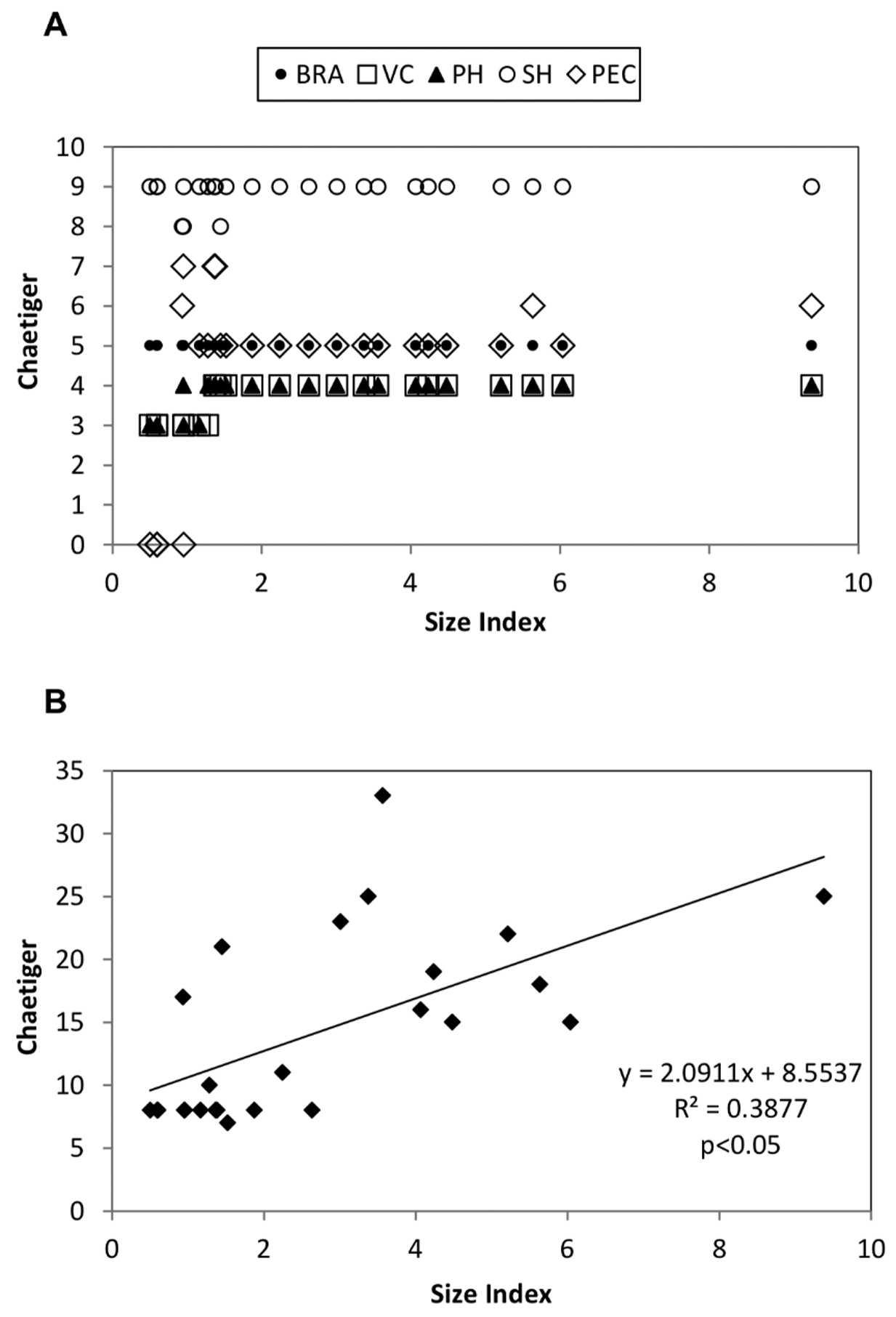
Intraspecific variability. The intraspecific variability of the main taxonomic characters of Onuphis farensis sp. nov. was analyzed in order to establish whether they are size related or subject to ontogenic variation, or if, contrary to that, are constant and reliable good diagnostic characters of the species (fig. 6). For the analyses the size of the worms was expressed through the Size Index as defined by Orensanz (1990) (see Material and Methods section).
The taxonomic characters analysed were: the first branchiate chaetiger (BRA), first chaetiger with pectinate chaetae (PEC), number of chaetigers with pseudocompound hooks ( PH), chaetigers with postchaetal lobes (PL), first chaetiger with subacicular hooks (SH), and number of chaetigers possessing ventral cirri (VC).
Figure 6 View FIGURE 6 A shows the results for all characters occurring in the first 9 chaetigers, i.e. BRA, PEC, PH, SH and VC. BRA was constant throughout the set of analysed specimens, always starting on chaetiger 5. SH showed almost no variation, starting in general on chaetiger 9 (with the exception of 3 juveniles discussed above, on which they first appear in only one of the parapodia of chaetiger 8, but are present in both parapodia of chaetiger 9). As in other smaller juveniles the SH was also chaetiger 9, these 3 exceptions represent probably anomalous specimens. PH and VC were normally present in the first 4 chaetigers, but in younger and smaller juveniles were only present in the first 3, which seems to represent an ontogenic variation. Finally, PEC is the character that showed greatest variation. Pectinate chaetae are normally present in adults from chaetiger 5, but in two adults they were first detected only in chaetiger 6. This could be due to: the absence of the chaetae in chaetiger 5, to its secondary loss in this chaetiger, or its presence being undetected. In very small juveniles however pectinate chaetae were absent, but when first present they appear first in chaetigers 6 or 7, and finally in chaetiger 5, a variation that seems to be related with the ontogeny of the species.
The relation between the number of chaetigers with postchaetal lobes (PL) and the Size Index is pictured in figure 6B. In this case the variability is larger, but there is a significant correlation (p <0.05) between the size of the worms as expressed by Orensanz’s Size Index and PL. Besides being size dependent, the bigger variability of this character can also be affected by the subjectivity of the observer deciding in which chaetiger they end, as postchaetal lobes decrease progressively in length before disappearing completely. Moreover, the fixation process and the different status in contraction of the specimens can also affect the shape of this structure. For this reason, the number of chaetigers with postchaetal lobes has a limited value, as a diagnostic character for the new species, and by extension, should also be avoided when characterizing other Onuphis species. The poor taxonomic value of the PL character was first suggested by Fauchald (1982b), and is herein confirmed.
Remarks. Onuphis farensis sp. nov. is the only species in the genus to possess both branchiae from chaetiger 5 and subacicular hooks from chaetiger 9 (see table 1). While some younger juveniles may not have pseudocompound hooks on the fourth chaetiger, this is a typical character of adults of this species. The presence of branchiae from chaetiger 5 and subacicular hooks from chaetiger 9 does not vary in juvenile specimens either, and therefore at the present date this is adequate to characterize the species. Three of the studied juveniles were aberrant by showing subacicular hooks on the 8th chaetiger, but only in one of the sides of the segment, being present in the two parapodia from chaetiger 9 onwards.
Onuphis farensis View in CoL sp. nov. was previously recorded and described by Ibáñez (1972), as Onuphis (Nothria) geophiliformis ( Moore, 1903) View in CoL , based on specimens from the sandbank of Cabezuela (Puerto Real, Bay of Cádiz, SW Spain), an environment similar to the Ria Formosa, but nowadays highly modified. The same author ( Ibáñez 1973a, 1973b) recorded O. geophiliformis View in CoL from Isla Cristina (Huelva, SW Spain), but without giving further details on the morphology of the specimens.
Onuphis geophiliformis View in CoL was originally described by Moore (1903, as Nothria View in CoL ), based on specimens collected north of Sendai Bay (Pacific Ocean off Japan), at 113 meters deep. Early posterior descriptions were provided first by Uschakov (1955, 1965), and later by Day (1960, 1967), in this case using newly collected material from South Africa. This resulted in a broadened definition of the species which fitted the specimens studied by Ibáñez (1972, from the Bay of Cádiz), particularly when using the available dicothomic keys from Uschakov (1965) and Day (1967). Fauchald (1982b) redescribed the species based on the type material, restricting its definition once more. Thus, it became evident that the populations from the Bay of Cádiz and Ria Formosa, and those from South Africa, TABLE 1. Synoptic table of valid species of the genus Onuphis View in CoL , showing selected morphological characters and type locality. Abbreviations: +, presence; ant., anterior; bi, bidentate; branch., branchiae or branchial; digit., digitiform; inter., interparapodial; max., maximum; no., number; papil., papilla; pect., pectinate; postset., postchaetal; pseud., pseudocompound; refs., references; set., chaetiger; spec., specimens; subac., subacicular; tetra, tetradentate; tri, tridentate). References: 1— Hilbig 1995; 2— Maekawa & Hayashi 1999; 3— Fauchald 1982b; 4— Fauchald 1982a; 5— Castelli 1982; 6— Ibáñez 1972; 7— Imajima 1986; 8— Maekawa & Hayashi 1989; 9— Shisko 1981; 10— Pettibone 1963; 11— Hobson 1971; 12— Claparède 1868; 13— Claparède 1869; 14— Claparède 1870; 15— Orensanz 1990; 16— Tan & Chou 1998; 17— Shen 1987.
Ant. set. w/ Ant. set. w/ digit. Set. w/ Teeth in pseud. hooks Branch. from set. Max. no. branch. Subac. hooks Start pect. set.
……continued on the next page TABLE 1. (Continued)
Ant. set. w/ Ant. set. w/ digit. Set. w/ pseud. Teeth in pseud. hooks Branch. from set. Max. no. branch. filaments Subac. hooks from Start pect. set. (no. vent. cirri postset. lobes hooks set. teeth)
longisetosa Imajima View in CoL , 5 about30 5 bi+tri 1 2 10 +(13–15)
(shorter from set. (in all set.) (up to set. 23–32, 1) 20)
mexicana View in CoL (Fauchald, 5 about13 5 tri 3–5 1 12–14 + (13–15)
) (usually 4) (usually 13)
multiannulata View in CoL 6 about 47 5 bi + tri 1 up to 3 10 + (13)
Shisko, 1981
nakaoi Maekawa View in CoL & 5 10 5 bi + tri 1 2 (2 after set. 35) 10 + (11)
Hayashi, 1999
nonpectinata View in CoL 4 10 3 tri 5 1 11 –12 notpresent
Imajima, 1986
opalina View in CoL (Verrill, 6 10–13 4–5 tri 1 1 12 –15 + (12)
) (range 5–7) (usually 13 or 14)
pallida View in CoL (Moore, 4 12 4 tri 4 1 16 +(15–20)
)
pancerii Claparède View in CoL ,?? 2 bi+tri 1 5–6 9 + (about 20)
(up to set. 16, 1)
pseudoiridescens View in CoL 4–6 11–12 3–4 tri 1 1 12 –15 +(12–14)
Averincev, 1972 (sometimes tetra) (sometimes 2–4)
punggolensis Tan View in CoL & 6 or 7 14 3 tri 1 4 10 4(10–11)
, 1998 (set. 1–16, 1; after, up to 4)
rullieriana View in CoL 5 or 6 12 4 tri 1 1 12 –14 +(15)
Amoureux, 1977) (3 in one
specimen)
shijikiensis Maekawa View in CoL 5 10+ 3 bi+tri 1 4 10 +(13)
Hayashi, 1999 (from set. 25)
shirikishinaiensis View in CoL 7 16–30 7 bi+tri 1 1 11 –14 +(16)
Imajima, 1960) (first 3 or 4 only tri; 4–7 (absent on last 40)
mainly bi)
similis View in CoL (Fauchald, 6 9 4 tri 6 1 8 or 9 + (10)
)
taraba Maekawa View in CoL & 5 11–12 4 tri 4 1 11 –13 +(16)
Hayashi, 1989
tetradentata View in CoL 4–5 10 4 tri+tetra 4–5 1 12 +(13–15)
Imajima, 1986 (set. 1–3, tetra; set. 4, tri)
texana Fauchald View in CoL , 6 10 2 bi 1 3 10 +(9)
(first 18–20, 1; next 10, 2; than, 2–3)
tosaensis Maekawa View in CoL 6 13 3 tri 1 4 10 +(10)
Hayashi, 1999 (4 from set. 19)
variolata Shen, 1987 View in CoL 6 10+ 8 bi + tri 1 up to 4 10 +(8)
(shorter from set. (4 at set. 19–30) 4–5)
vibex View in CoL (Fauchald, 6 8 or 9 5 tri 4 1 13 +(10)
) (lowermost tooth of some
hooks furcate)
wakasaensis 5 14 4 tri+tetra 4 1 13 +(18)
Maekawa & Hayashi, (set. 2, 1 tri + 2 tetra; set.
3, 3 tetra; set. 4, 1 tetra)
……continued on the next page Inter. papil. Maxillary formula Tube Type locality Refs.. affinis Hilbig, 1995 View in CoL ? MI: 1+1, MII: 7–9+8–10, MIII: 9+0, MIV:? Central California, Pacific Ocean, 985 m 1
7–8+11, MV: 1+1
. amasukaensis Maekawa & Hayashi ,? MI: 1+1, MII: 7+7, MIII: 7+0, MIV: 5+8,? Japan, East China Sea, intertidal 2 1999 MV: 1+1
. aucklandiensis Augener, 1924 ??? off New Zealand, Pacific Ocean, 27 m 3. branchiata Treadwell, 1931 View in CoL ? MI: 1+1, MII: 8+9, MIII: 8+0, MIV: 6+6,? Philippine Islands, Pacific Ocean, 193 m 3
MV: 1+1
. chinensis Uschakov & Wu, 1962 View in CoL no MI: 1+1, MII: 8+9, MIII: 7+0 MIV: 7+10,? YellowSea 3
MV: 1+1
. declivorum Fauchald, 1982 View in CoL ??? off New Jersey, Atlantic Ocean, 350 m 4. elegans ( Johnson, 1901) View in CoL ? MI: 1+1, MII: 6-7+5-7, MIII: 7-10+0,? West coast of USA and British Columbia, 3
MIV: 6-7+5-8, MV: 1+1 Canada, shallow water
. eremita Audouin & Milne-Edwards View in CoL , 4–10 MI: 1+1, MII: 6-7+8-9, MIII: 8-9+0, MIV: cylindrical, thin walled, covered with La Rochelle, Atlantic Ocean, shallow 3 1833 4 +9-10, MV: 1+1 sand grains water. eremita oculata Hartman, 1951 View in CoL 4–10 MI: 1+1, MII: 8+9, MIII: 9+0, MIV:? Florida, Gulf of Mexico, intertidal to 3
6+11, MV: 1+1 shallow waters
. eremita parva Berkeley & Berkeley View in CoL , 4–10?? Central California, 22–31 m 3 1941
. falesia Castelli, 1982 View in CoL ? MI: 1+1, MII: 8-9+8-9, MIII: 8+0, MIV: membranous encrusted with sand Gulf of Follonia, Italy, infralittoral 5
6-8+10, MV: 1+1 grains, small stones, shell fragments
. farallonensis Hobson, 1971 View in CoL ? MI: 1+1, MII: 9+10, MIII: 10+0, MIV:? off Northern California, Pacific Ocean, 67 3
6+11, MV: 1+ 1 m
. farensis Gil & Machado View in CoL , new species no MI: 1+1, MII: 9+9, MIII: 10+0, MIV: membranous encrusted with sand Ria Formosa, Portugal; intertidal 6, this
7+10, MV: 1+1 grains of different sizes work. fuscata Imajima, 1986 View in CoL ? MI: 1+1, MII: 9+9, MIII: 8+0, MIV:? NE Japan, Pacific Ocean, 40–44 m 7
10+12, MV: 1+1
. geophiliformis ( Moore, 1903) View in CoL ? MI: 1+1, MII: 9+9, MIII: 8+0, MIV: 7+9,? off Japan, Pacific Ocean, 113 m 2, 3
MV: 1+1
. hokkaiensis Maekawa & Hayashi View in CoL ,? MI: 1+1, MII: 7+8, MIII: 8+0, MIV: 6+7,? Japan, Pacific Ocean, depth unknown 2 1999 MV: 1+1
. holobranchiata Marenzeller, 1879 View in CoL ? MI: 1+1, MII: 6+6-8, MIII: 7+0, MIV: 5-? off Japan, Pacific Ocean, depth unkown 2, 3
7+9-12, MV: 1+1
. imajimai Maekawa & Hayashi, 1989 View in CoL ? MI: 1+1, MII: 7+8, MIII: 8+0, MIV: 6-? Japan, Sea of Japan, 150–295 m 2, 8
8+9-10, MV: 1+1
. iridescens ( Johnson, 1901) View in CoL ? MI: 1+1, MII: 7+8, MIII: 8+0, MIV: inner lining thick and stiff, sparsely off British Columbia, Pacific Ocean 3
5+10, MV: 1+1 covered with sand grains
. iriei Maekawa & Hayshi, 1999 View in CoL ? MI: 1+1, MII: 9+9, MIII: 9+0, MIV: 5+8, semitransparent membrane southern East China Sea, 82 m 2
MV: 1+1
. kammurijimaensis Maekawa View in CoL &? MI: 1+1, MII: 7+9, MIII: 10+0, MIV:? Japan, Sea of Japan, 72 m 8 Hayashi, 1989 7+10, MV: 1+1
……continued on the next page represented undescribed taxa (see table 2), distinct from O. geophiliformis View in CoL . This theory appears supported by the different geographical origins of the populations. The population from the Southern Iberian Peninsula is herein described as Onuphis farensis View in CoL sp. nov., whilst the status of the South African population requires further investigation, in order to confirm its identity as a second new species.
Onuphis farensis View in CoL sp. nov. was collected at the type locality between the 25th November 1999 and the 12th June 2003, but a collecting trip to the type locality performed on the 17th June 2014 did not provide any additional specimens. The Ria Formosa coastal lagoon is a dynamic system, and the intertidal sandbank where the species was initially found is currently covered by a meadow of Zostera noltii Hornemann View in CoL , which is being replaced again by sand. The periodical changes in the type locality, as well as the possible patchy distribution of the species, are possible explanations for the absence of specimens in the new samples.
Locality Senday Bay (Off Japan) Cape ( South Africa) Cádiz ( Spain) Faro ( Portugal)
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Onuphis farensis
| Gil, João & Machado, Margarida 2014 |
Onuphis
| Gil 2011: 544 |
Onuphys geophyliformis
| Mucha 1999: 371 |
Nothria geophiliformis
| Dexter 1992: 79 |
Nothria geophiliformis
| Campoy 1982: 557 |
Onuphis (Nothria) geophiliformis
| Ibanez 1973: 128 |
| Ibanez 1973: 43 |
| Ibanez 1972: 26 |