
Aeolosoma major
Rouse, Greg W. & Pleijel, Fredrik, 2007, Annelida *, Zootaxa 1668, pp. 245-264 : 250-255
|
publication ID |
https://doi.org/ 10.5281/zenodo.180168 |
|
DOI |
https://doi.org/10.5281/zenodo.6244305 |
|
persistent identifier |
https://treatment.plazi.org/id/03FC87EB-FFA0-5820-FF45-FE4BF4BEF93F |
|
treatment provided by |
Plazi |
|
scientific name |
Aeolosoma major |
| status |
|
Major polychaete taxa
Scolecida. The name Scolecida, derived from skolex (Greek), meaning worm, was applied with reference to an annelid group by Rouse & Fauchald (1997), though it has historically been used with reference to parasitic non-annelids. Taxa included were Arenicolidae , Capitellidae ( Fig. 1 C inset), Cossuridae , Maldanidae ( Fig. 2 View FIGURE 2 E), Opheliidae ( Fig. 2 View FIGURE 2 D), Orbiniidae ( Fig. 2 View FIGURE 2 C) and Paraonidae . The name Scolecida is derived from Scoleciformia, a name introduced by Benham (1896) for a similar group of taxa. Only two apomorphies support the clade Scolecida in Rouse & Fauchald (1997), the presence of parapodia with similar rami and the possession of two or more pairs of pygidial cirri, and both of these are homoplastic. In many ways, this group represents the simple-bodied forms of polychaetes and it is likely that further analysis will show that it is not monophyletic. In fact to date no molecular sequence analyses have recovered any assemblage like Scolecida and it usually appears as a polyphyletic assemblage (e.g., Struck et al. 2007).
Palpata. Palpata was a new name coined by Rouse & Fauchald (1997) and such a group of polychaetes has never been formulated before. Virtually all non-Scolecida polychaetes, except a few incertae sedis taxa, were placed into Palpata. The name is based on an apomorphy for the group, the presence of palps. Palps can be divided into two structurally different groups, grooved “feeding” palps and ventral, “sensory” palps ( Orrhage 1980). Feeding palps usually have ciliated paths, often located in a longitudinal groove, giving each palp a Ushaped cross-section. Ventral sensory palps are morphologically more uniform than grooved palps. In most cases they are tapering or digitiform and relatively short, compared to grooved palps. Given the fact that the presence of palps and a limited peristomium were the only synapomorphies supporting this taxon in Rouse & Fauchald (1997), the validity of Palpata requires further investigation and it has yet to be recovered as a monophyletic group in any broad molecular analyses (see below). Rouse & Fauchald (1997) proposed that Palpata contains two major taxa, Aciculata and Canalipalpata .
Aciculata. Aciculata was a new name erected by Rouse & Fauchald (1997) for one of the most the strongly supported clades in their analyses. The name refers to one of the apomorphies for the group; the presence of a particular chaetal type called aciculae. Aciculae are stout chaetae that differ from others in that much or all of the chaeta remains internalized in the parapodium. Numerous other features such as the presence of ventral sensory palps, prostomial antennae, dorsal cirri, ventral cirri, one pair of pygidial cirri, and segmental organs in most segments were proposed to be apomorphic for Aciculata. Aciculata is divided here into the three major clades Amphinomida , Eunicida and Phyllodocida , and also with a few taxa as incertae sedis. Aciculata was recovered as a clade by Struck et al. (2007), if Orbiniidae (usually placed in Scolecida, although some members actually have chaetae that can be interpreted as aciculae) was included.
Phyllodocida View in CoL . The name Phyllodocida View in CoL was coined by Dales (1962) and was subsequently used by other workers, such as Fauchald (1977) and Pettibone (1982), for essentially the same group of polychaetes. Prior to Dales (1962), the group had never been formulated as a monophyletic taxon. Rouse & Fauchald (1997) found strong support for the monophyly of this assemblage, indicated by the ventral position of sensory palps, the presence of anterior enlarged cirri, the loss of dorsolateral folds, the presence of an axial muscular proboscis, and the presence of compound chaetae with a single ligament. A major difference in the formulation of Phyllodocida View in CoL by Rouse & Fauchald (1997), and that of previous workers, was that they placed Myzostomida ( Fig. 1 D, E) (as Myzostomatidae ) in the group (but see Eeckhaut et al., 2000). Rouse & Fauchald (1997) did not further subdivide Phyllodocida View in CoL , since their analyses resulted in dramatically different topologies for various Phyllodocida View in CoL taxa. Based on the results of Pleijel & Dahlgren (1998), two names were used by Rouse & Pleijel (2001) for clades within Phyllodocida View in CoL ; Aphroditiformia (or scale-worms) and Nereidiformia. The remaining taxa in Phyllodocida View in CoL are treated as “unplaced”. There are numerous family-ranked taxa in Phyllodocida View in CoL with representatives shown here of Phyllodocidae View in CoL ( Fig. 3 A), Sphaerodoridae View in CoL ( Fig. 3 B), Nereididae View in CoL ( Fig. 3 C), Syllidae View in CoL ( Fig. 3 D), Chrysopetalidae View in CoL ( Fig. 4 A), Nephtyidae View in CoL ( Fig. 4 B), and Polynoidae View in CoL ( Fig. 4 E).
Eunicida View in CoL . The name Eunicida View in CoL was first used by Dales (1962), but encompassed a well-delineated group known prior to that as Eunicea, Euniceformia, Eunicimorpha or simply Eunicidae View in CoL . This grouping included polychaetes with a ventral muscularized pharynx with ventral mandibles and dorsal maxillae (and a few that had lost jaws) and is very likely a monophyletic assemblage. Another putative apomorphy for the clade would appear to be a peristomium forming a two rings, although this feature is not present in all the subgroups. Rouse & Fauchald (1997) expanded the traditionally delineated Eunicida View in CoL to include Amphinomidae View in CoL and Euphrosinidae View in CoL although this is not accepted here as the relationships between them is uncertain. Eunicida View in CoL , following Rouse & Pleijel (2001), includes Dorvilleidae View in CoL ( Fig. 4 C), Eunicidae View in CoL , Hartmaniellidae View in CoL , Histriobdellidae View in CoL , Lumbrineridae View in CoL , Oenonidae View in CoL and Onuphidae View in CoL . A recent comprehensive molecular phylogeny of Eunicida View in CoL was undertaken by Struck et al. (2006) who found a topology where a labidognath clade (i.e., a “ Eunicidae View in CoL ”/ Onuphidae View in CoL / Lumbrineridae View in CoL clade) was significantly rejected, a result that goes against most traditional view of the relationships within the group. Prior to this Struck et al. (2005), in a molecular study suggested that Dinophilidae View in CoL ( Fig. 1 A)—a group long associated with Eunicida View in CoL , either as a family or as part of Dorvilleidae ( Eibye-Jacobsen & Kristensen 1994) View in CoL —does not belong in Eunicida View in CoL .
Amphinomida View in CoL . Amphinomida View in CoL contains Amphinomidae View in CoL ( Fig. 4 D) and Euphrosinidae View in CoL and has never been the subject of a detailed modern phylogenetic revision. A taxon, Amphinomae, equivalent to Amphinomida View in CoL , dates back to Savigny (1822). Amphinomida View in CoL was first used as a taxon name by Dales (1962), but names such as Amphinomorpha have also been applied. Although Amphinomida View in CoL is well supported by a number of apomorphies, including calcareous chaetae and nuchal organs forming a caruncle, both their position within Aciculata and the interrelationships within the group are uncertain. Rouse & Fauchald (1997) in their classification included Amphinomidae View in CoL and Euphrosinidae View in CoL in Eunicida View in CoL , whereas Rouse & Pleijel (2001) instead treated them as a separate taxon, Amphinomida View in CoL .
Aciculata unplaced. Rouse & Fauchald (1997) placed Aberranta Hartman, 1965 View in CoL (as Aberrantidae View in CoL ), Nerillidae View in CoL ( Fig. 1 H) and Spinther Johnston, 1845 View in CoL (as Spintheridae View in CoL ) as parts of Aciculata, but did not include them in any other sub-taxon of the group. These three taxa have never been related to each other and are not to be regarded as closely related by being grouped together here. Of these taxa, only Spinther View in CoL actually has aciculae, but all three are likely to have sister groups with different members of Aciculata. Worsaae et al. (2005) assessed the phylogenetic position of Nerillidae View in CoL and Aberranta View in CoL using morphology and 18S rDNA, but failed to provide any strong conclusions as to the placement for either group.
Canalipalpata . Canalipalpata (referring to the presence of grooved palps) is a name that was first used by Rouse & Fauchald (1997) and is a massive group of polychaetes that encompasses around half the number of described species. The only apomorphy for Canalipalpata is the presence of grooved palps and so the clade must be regarded as weakly supported. The groove along each palp is longitudinal and ciliated and, in contrast to those of Aciculata, function as feeding structures. Rouse & Fauchald (1997) identified three major clades within the Canalipalpata ( Sabellida , Spionida and Terebellida ) and a number of taxa that were regarded as incertae sedis within the group.
Sabellida View in CoL . The apomorphy for Sabellida View in CoL , as formulated by Rouse & Fauchald (1997), was the fusion of the prostomium with the peristomium, hence it is weakly supported. This name has been used to contain Sabellariidae View in CoL ( Fig. 5 I), Sabellidae View in CoL ( Fig. 5 C) and Serpulidae ( Fitzhugh, 1989) View in CoL and these three were included by Rouse & Fauchald (1997), plus two other taxa. A dramatic shift compared with traditional systematics was that Rouse & Fauchald (1997) placed Siboglinidae View in CoL ( Fig. 5 A), (formerly outside polychaetes as Pogonophora and Vestimentifera), as part of Sabellida View in CoL . They also included Oweniidae View in CoL ( Fig. 5 B), a taxon that has a “chequered” systematic history as a polychaete. It should be noted that in further analyses by Rouse (1999; 2000), Oweniidae View in CoL did not group with the remaining Sabellida View in CoL , whereas Chaetopteridae View in CoL ( Fig. 5 F) (included here in as part of Spionida View in CoL ) did. A recent combined analysis of molecular and morphological data ( Rousset et al. 2004) also did not recover the Sabellida View in CoL as formulated here but did suggest Oweniidae View in CoL and Siboglinidae View in CoL to be closely related, as did Struck et al. (2007). Clearly further study is required. In addition, similarities between Sabellariidae View in CoL ( Fig. 5 I) and Pectinariidae View in CoL (in Terebellida View in CoL ) also deserve further study ( Rouse & Pleijel 2001). Sabellariidae View in CoL have previously been considered as part of Terebellida View in CoL (e.g., Fauchald 1977).
Terebellida (Cirratuliformia, Terebelliformia) View in CoL . The name Terebellida View in CoL was first used by Dales (1962) and included Ampharetidae View in CoL , Pectinariidae View in CoL and Terebellidae View in CoL , all polychaetes having multiple grooved palps. Prior to this, the names Terebellomorpha or Terebelliformia had often been used for essentially the same grouping of taxa. Terebellida View in CoL was expanded by Rouse & Fauchald (1997) to include a clade in which most have a single pair of palps (e.g., Acrocirridae View in CoL , Cirratulidae View in CoL , Flabelligeridae View in CoL ), but recent studies have not recovered this relationship (see below). Rouse & Fauchald (1997) identified several synapomorphies for this overall grouping, namely the presence of a first segment with no chaetae, a gular membrane, and a heart body. The clade within Terebellida View in CoL that has taxa with a single pair of palps (with exceptions such as some Cirratulidae View in CoL with numerous palps and Ctenodrilinae and Fauveliopsidae View in CoL with none) was referred to as Cirratuliformia in Rouse & Pleijel (2001), a name that has been used previously for a somewhat similar grouping (e.g., Fauchald 1977). It contains Acrocirridae View in CoL , Cirratulidae View in CoL , Fauveliopsidae View in CoL , Flabelligeridae View in CoL , and Sternaspis Otto, 1821 View in CoL . The clade comprised of Alvinellidae View in CoL , Ampharetidae View in CoL , Pectinariidae View in CoL , Terebellidae View in CoL and Trichobranchidae View in CoL is referred to here as Terebelliformia, also a fairly “traditional” formulation.
Spionida View in CoL . The name Spionida View in CoL was first used by Dales (1962) to contain Spionidae View in CoL and a number of similar groups, as well as taxa such as Paraonidae View in CoL and Sabellariidae View in CoL . These groups are not considered closely related today, and Spionida View in CoL in Rouse & Pleijel (2001) contained Apistobranchus Levinsen, 1883 View in CoL , Chaetopteridae View in CoL , Magelonidae View in CoL , Heterospio Ehlers, 1875 View in CoL , Poecilochaetus Claparède View in CoL in Ehlers, 1875, Trochochaeta Levinsen, 1883 View in CoL , and Uncispionidae View in CoL , though the latter four taxa should reasonably be all treated as members of Spionidae ( Blake & Arnofsky 1999) View in CoL . The positions of Chaetopteridae View in CoL and Magelonidae View in CoL deserve further investigation. The synapomorphies for Spionida View in CoL listed by Rouse & Fauchald (1997) were the presence of a pair of peristomial grooved palps, nuchal organs forming posterior projections, and anterior excretory nephridia and posterior segmental organs for gamete release.
Canalipalpata unplaced. Rouse & Fauchald (1997) placed Polygordiidae View in CoL and Protodrilida (as Protodriloididae View in CoL , Protodrilidae View in CoL and Saccocirridae View in CoL ) as part of Canalipalpata , but did not place them within any other subtaxon. In their complete cladistic analyses, these taxa formed a clade that was either associated with taxa that belong within Canalipalpata, Scolecida or were part of a basal polytomy of Polychaeta, so their decision was arbitrary. However, there is good evidence to support Protodrilida being placed in Canalipalpata ( Purschke & Jouin 1988) , perhaps near Spionida View in CoL . On the other hand, placement of Polygordiidae View in CoL in Canalipalpata must be regarded as suspect. Rouse & Fauchald (1997) made scoring errors in regard to Polygordiidae View in CoL , the most important being that the animals cannot be regarded as having grooved palps. It was suggested by Rouse & Pleijel (2001) that investigation of a sister group relationship with, or within, Opheliidae View in CoL may be worthwhile, and this is actually an old idea (e.g., Giard 1880).
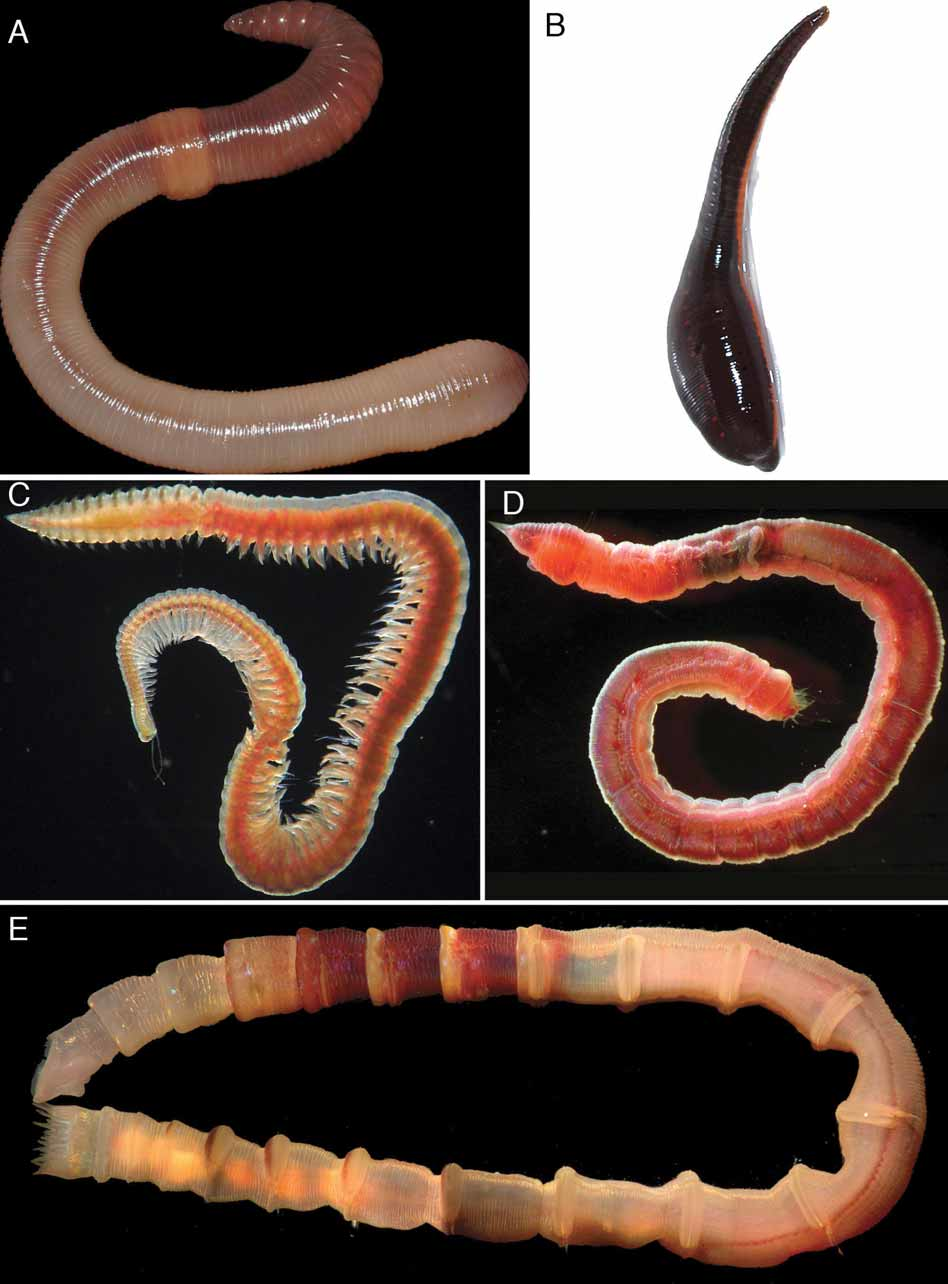
Annelida unplaced. Rouse & Fauchald (1997) included Aeolosomatidae ( Fig. 1 B), Potamodrilus Lastockin, 1935 (as Potamodrilidae ), Parergodrilidae View in CoL and Psammodrilidae View in CoL in their complete taxon set analyses. In one analysis, these taxa either fell as part of a large basal polytomy of polychaetes, or Aeolosomatidae , Potamodrilus and Parergodrilidae View in CoL formed a clade with Ctenodrilinae , and Psammodrilidae View in CoL grouped with Capitellidae View in CoL ( Fig. 1 C inset), Arenicolidae View in CoL and Maldanidae View in CoL ( Fig. 2 View FIGURE 2 E). In another analysis, Aeolosomatidae , Potamodrilus and Parergodrilidae View in CoL either formed a grade with respect to Polygordiidae View in CoL ( Fig. 1 F), Protodrilidae View in CoL ( Fig. 1 G) and Protodriloididae View in CoL , or they were a basal clade of polychaetes. Psammodrilidae View in CoL either were a basal polychaete group, or were sister group to a large clade that was mainly comprised of taxa with grooved palps. Thus, it was not possible to place these taxa with any confidence. Prior to the study by Rouse & Fauchald (1997), Aeolosomatidae ( Fig. 1 B) and Potamodrilus were usually considered in relation to Clitellata, either as a member of the group ( Bunke 1967), or more recently as the sister group to Clitellata ( Bunke 1985). This was then rejected by Bunke (1986), who also could not relate them to any group of polychaetes, thus leaving the two taxa “isolated”. The most recent analysis is from a molecular perspective ( Struck et al. 2002), and this study found no support for a relationship of Aeolosomatidae with Clitellata or with any other particular annelid group. Fauchald (1977) grouped Parergodrilidae View in CoL with Ctenodrilidae , but did not justify this decision. Otherwise, the group has also been treated as “isolated” within polychaetes. A recent molecular study by Jördens et al. (2004) indicated that Parergodrilidae View in CoL may be closely related to orbiniids; the position of the terrestrial polychaete Hrabeiella Pizl & Chalupsky, 1984 , however, was inconclusive. Psammodrilidae View in CoL have been treated as a singular group of polychaetes since they were first discovered by Swedmark (1952). Rouse & Fauchald (1997) suggested that a relationship for Psammodrilidae View in CoL with Arenicolidae View in CoL and Maldanidae View in CoL should be assessed; this was proposed by Meyer & Bartolomaeus (1996; 1997), but the present anatomical evidence is weak.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Aeolosoma major
| Rouse, Greg W. & Pleijel, Fredrik 2007 |
Spionidae (
| Blake & Arnofsky 1999 |
Dorvilleidae (
| Eibye-Jacobsen & Kristensen 1994 |
Serpulidae (
| Fitzhugh 1989 |
Canalipalpata (
| Purschke & Jouin 1988 |
Hrabeiella
| Pizl & Chalupsky 1984 |
Aberranta
| Hartman 1965 |
Potamodrilus
| Lastockin 1935 |
Sphaerodoropsis philippi
| Fauvel 1911 |
Apistobranchus
| Levinsen 1883 |
Trochochaeta
| Levinsen 1883 |
Bylgides elegans (Théel, 1879)
| Theel 1879 |
Heterospio
| Ehlers 1875 |
Eulalia clavigera
| Quatrefages 1866 |
Spinther
| Johnston 1845 |
Sternaspis
| Otto 1821 |
Nephtys hombergi
| Savigny 1818 |
Hediste diversicolor (O.F. Müller, 1776) (Nereididae)
| O.F. Muller 1776 |